Necessidades Térmicas e Fotoperiódicas na Cultura do Milho


A adaptação climática de qualquer espécie vegetal tem estreita relação com seu padrão fenológico. Dela depende a distribuição geográfica de cada espécie e sua flutuação nos ecossistemas. Em cultivos agrícolas, o padrão fenológico é requisito fundamental para o zoneamento de cada espécie, visando indicação de regiões e épocas preferenciais. Para uma mesma espécie, o zoneamento se torna mais preciso ao considerar a variabilidade fenológica entre genótipos, que podem ser classificados por grupos de maturação.
Este processo permite adequar as necessidades das plantas às disponibilidades do ambiente, em particular às condições climáticas. Neste caso, não se trata simplesmente da ocorrência, frequência e sobrevivência de indivíduos em diferentes ambientes. Trata-se, sobretudo, de explorar a potencialidade dos cultivos e manter a sustentabilidade dos sistemas de produção.
A temperatura do ar é um dos principais fatores determinantes da fenologia das plantas. Por isto, ela exerce papel fundamental na distribuição espaço-temporal das espécies. A interação das condições térmicas com a duração dos dias (fotoperíodo) também é considerada importante para inúmeras espécies cultivadas. O fotoperíodo exerce influência no ciclo de muitas espécies e este efeito é denominado fotoperiodismo.
Em cultivos agrícolas o maior interesse do fotoperiodismo decorre, sobretudo, da indução ao florescimento, que interfere no crescimento das plantas e na duração do ciclo, para muitas espécies. Em diversas culturas (como a soja) o fotoperíodo pode ser fator determinante da distribuição regional de cultivares e do próprio calendário de cultivo.
Em milho o início do período reprodutivo se dá quando o meristema apical começa a se alongar e inicia o primórdio das flores do pendão (masculinas). Como nenhuma folha poderá formar-se após a diferenciação do meristema apical, a iniciação do primórdio floral determina o número máximo de folhas, nós e internos de cada colmo e, portanto, o tempo relativo entre a emergência e o final do período vegetativo. O número de nós formados antes da iniciação do pendão (portanto, o número de folhas) depende do genótipo, mas é modificado pela temperatura e o fotoperíodo (DUNCAN, 1976).
Segundo Chang (1974) a espécie Zea mays pode ser considerada fotoneutra (isto é, sem resposta ao fotoperíodo) ou de resposta a dias curtos. Ou seja, dependendo dos genótipos e das condições regionais (latitude e época), a fenologia do milho pode ser influenciada pelo fotoperíodo. Neste caso, em genótipos que respondem a dias curtos o encurtamento do fotoperíodo tende a reduzir as necessidades térmicas para a indução ao pendoamento.
Em experimentos na Austrália, Birch, Hammer e Rickert (1998) verificaram que o aumento do fotoperíodo pode prolongar o crescimento vegetativo (da emergência ao pendoamento) e aumentar o número de folhas. Este efeito se manifestou em genótipos adaptados ao trópico, quando foram semeados no final de verão daquele País.
Por outro lado, Birch, Vos e Van der Putten, (2003) testaram genótipos de diferentes ciclos em maior latitude (na Holanda), incluindo um genótipo adaptado ao subtrópico australiano, e confirmaram que o milho tem pequena ou nenhuma resposta ao fotoperíodo.
Estes estudos demonstraram que a evolução da fenologia e formação de órgãos em plantas de milho (folhas, em particular) é função do tempo térmico, expresso por graus-dia acima de 8ºC. O genótipo australiano manifestou a mesma necessidade térmica na Holanda e na Austrália, com fotoperíodo entre 12,5 e 16,5h. Houve diferenças no tempo térmico entre genótipos, por serem de diferentes grupos de maturação.
Também houve maior necessidade de graus-dia nas primeiras épocas de semeadura, as quais foram atribuídas às baixas temperaturas ocorridas, em muitos casos abaixo da temperatura mínima basal, causando estresses por resfriamento (chilling) e retardando o desenvolvimento das plantas.
Sabe-se, porém, que a fenologia do milho é regulada basicamente pelo regime térmico, o que permite o emprego de modelos de crescimento baseados em somas térmicas. Inúmeros trabalhos demonstraram que os genótipos de milho cultivados nas diferentes regiões do Brasil necessitam determinada quantidade de energia para cumprir suas etapas fenológicas e o próprio ciclo.
Assim sendo, para fins práticos e embora possa haver influências do fotoperíodo, o milho tem sido considerado uma espécie sem resposta ou de pouca resposta fotoperiódica. Portanto, o desenvolvimento da planta do milho depende, basicamente, das condições térmicas.
O uso da soma de graus-dia, baseada no acúmulo energético acima de determinada temperatura-base, é de uso abrangente em modelos que descrevem o desenvolvimento fenológico e o crescimento do milho. Dentro de um mesmo grupo de maturação, é possível estimar a ocorrência de fases da cultura, para diferentes genótipos, regiões e épocas de cultivo, utilizando a temperatura do ar como única variável.
Embora a radiação solar e a água também possam influenciar a fenologia do milho, a soma de graus-dia (temperatura do ar, portanto) tem relação linear com o desenvolvimento das plantas de milho (COELHO; DALE, 1980). A interação com outros fatores, como água, nitrogênio e o próprio fotoperíodo, pode provocar pequenas margens de erro, sob diferentes ambientes, mas o somatório de graus-dia tem sido um adequado parâmetro para estimar eventos fenológicos de milho.
No trabalho clássico de Gilmore e Rogers (1958) o número de unidades de calor para o espigamento, acima de 10ºC (designadas pelo autor como “graus efetivos”), permaneceu relativamente constante para diferentes épocas, enquanto os dias do calendário variaram amplamente.
A partir daí, o critério de “tempo térmico” se tornou de uso crescente, substituindo o tempo cronológico (dias do calendário) pela redução na variabilidade entre ambientes diferentes. A predição de períodos críticos, datas da floração e maturação, a programação de práticas de manejo importantes e o emprego generalizado da modelagem são alguns exemplos de aplicações dos graus-dia em milho (DAUGHTRY; COCHRAN; HOLLINGER, 1984).
As condições térmicas influenciam os mais diversos processos vitais das plantas, desde germinação e emergência pela temperatura do solo, o desenvolvimento fenológico e o crescimento da planta como um todo, por ambas as temperaturas do solo e ar. Durante o período de crescimento vegetativo, o tempo térmico decorrido entre o aparecimento de duas folhas sucessivas é denominado “filocrono”.
Este índice permite estimar a duração do período vegetativo das plantas e, portanto, a época de florescimento em função da temperatura do ar. No Estado de São Paulo, Tojo Soler, Sentelhas e Hoogenboom (2005) determinaram o filocrono de diversos híbridos de milho, considerando duas temperaturas-base: 8ºC e 10ºC.
Verificaram que o aparecimento de cada folha necessitou cerca de 50 graus-dia até a décima segunda folha, diminuindo após. Isto demonstra que é possível fazer uma simulação precisa da fenologia do milho e da evolução do índice de área foliar no período vegetativo a partir do tempo térmico, devido à influência direta da temperatura no metabolismo das plantas e à alta correlação entre o número final de folhas e a duração do período vegetativo.
A lei de Van´t Hoff aplicada ao crescimento vegetal sugere que a velocidade dos processos metabólicos duplica a cada 10°C de aumento da temperatura. Porém, não é correto considerar que as plantas mantenham este padrão de resposta indefinidamente.
Assim, o efeito da elevação da temperatura sobre o metabolismo das plantas não se expressa através de uma função linear contínua. Alguns estudos verificaram que modelos não lineares têm melhor desempenho em simular a fenologia do milho, tanto no período vegetativo como no reprodutivo. (BONHOMME, 2000; GUISCEM et al., 2001; STRECK et al., 2008).
O método de graus-dia em milho apresenta outras limitações, como o efeito diferenciado da temperatura do ar ao longo do ciclo da cultura e diferenças de necessidades térmicas entre genótipos, sobretudo de temperaturas-base (DIDONET et al., 2008a, 2008b).
Entretanto, mesmo admitindo as imperfeições do método e os possíveis efeitos das condições hídricas e do fotoperíodo, modelos lineares associando a fenologia do milho ao acúmulo de graus-dia tem sido amplamente testados e utilizados no Brasil (BERLATO; MATZENAUER; SUTILI, 1984; LOZADA; ANGELOCCI, 1999; GADIOLI et al., 2000; TOJO SOLER; SENTELHAS; HOOGENBOOM, 2005).
A Figura 6 demonstra, de uma forma genérica, como a temperatura influencia o metabolismo básico das plantas (BERGONCI; BERGAMASCHI, 2002). A fotossíntese líquida (assimilação líquida) tem aumento acentuado em baixas temperaturas, ocasionado por incrementos diferenciados nos processos de fotossíntese e respiração, atingindo um máximo na chamada temperatura ótima.
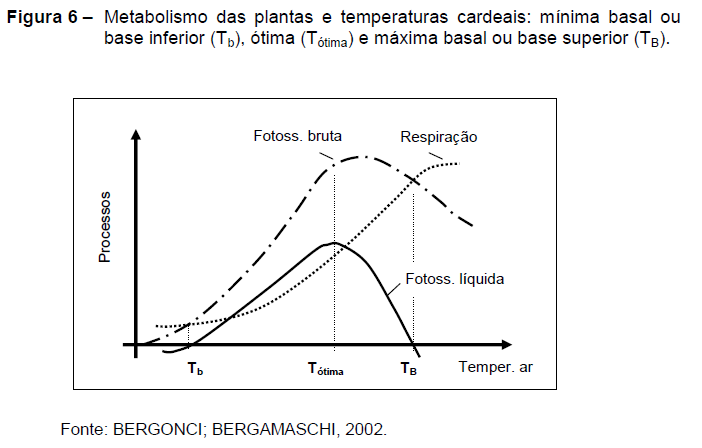
Em temperaturas acima da ótima a assimilação líquida diminui, já que os gastos por respiração aumentam mais do que os ganhos pela fotossíntese. A fotossíntese líquida é positiva entre os dois extremos de adaptação da espécie ou cultivar, chamados temperatura base inferior (Tb) e temperatura base superior (TB). Fora desses dois limites a assimilação líquida é negativa, sendo que as plantas paralisam seu crescimento e passam a sofrer estresse térmico.
A interpretação da Figura 6 serve como base para as relações de causa-efeito entre a temperatura do ar e o desenvolvimento das plantas (crescimento e fenologia). No período de germinação das sementes e emergência das plântulas a lógica é semelhante com relação à temperatura do solo, sendo que cada espécie tem seus limites de tolerância e uma faixa térmica ótima.
O cálculo do acúmulo de graus-dia toma por base os limites térmicos de cada espécie ou genótipo, que são as chamadas temperaturas-base: inferior (Tb) e superior (TB). Rigorosamente, o conceito de graus-dia deveria considerar ambos os limites térmicos e desprezar os períodos em que a temperatura estiver abaixo de Tb e acima de TB.
Entretanto, na grande maioria dos trabalhos e modelos, o cálculo do tempo térmico leva em consideração somente o limite inferior (Tb), efetuando a soma de “temperaturas efetivas” (como alguns autores preferem), ou seja, a diferença entre a temperatura-base inferior e a temperatura média diária.
As temperaturas cardeais (ótima, base inferior e base superior) são mais elevadas em plantas de clima quente do que nas de clima frio. Dentro de uma mesma espécie, cultivares precoces têm suas temperaturas cardeais mais baixas que cultivares tardias. Esta diferença de resposta, adquirida através do melhoramento genético, permitiu que o milho pudesse ser cultivado em regiões de climas tão diferenciados.
Também, tornou-se viável cultivar milho em regiões que eram consideradas inaptas por insuficiência térmica, como em planaltos das regiões Sul e Sudeste do Brasil. Do mesmo modo, o surgimento de genótipos mais precoces tornou possível a antecipação de épocas de cultivo no Sul do Brasil, assim como o cultivo em safrinha, ao final do verão e início de outono, sob temperaturas relativamente baixas.
Nas estimativas do acúmulo de graus-dia, é importante considerar as diferenças nas temperaturas cardeais entre os grupos de maturação. Diversos autores obtiveram melhores estimativas fazendo variar esses índices, de acordo com o tipo de genótipo. Berlato e Sutili (1976) obtiveram como melhores temperaturas mínimas basais 4°C em híbridos precoces, 6°C para os de ciclo médio e 8°C para tardios, pela classificação adotada na época.
Porém, Kiniry (1991) considerou como limites extremos para a fenologia do milho 8°C e 44°C, sendo que o crescimento máximo ocorre entre 26 e 34°C. Para diversos genótipos da Austrália, Birch, Hammer e Rickert (1998) adotaram as seguintes temperaturas cardeais para cálculo de graus-dia: Tb = 8ºC, Tótima = 30ºC e TB = 40ºC.
A classificação pela precocidade de genótipos através de denominações do tipo precoce, superprecoce, tardio, etc. é imprecisa para uso em grande escala. Ela depende de características genéticas, porém as interações genótipo-ambiente se alteram sempre que as condições do meio se modificam. Portanto, ela é relativa às condições do ambiente e, no caso particular do milho, as condições térmicas são aquelas que mais influenciam a duração das etapas fenológicas.
Por este motivo, o ciclo de cada genótipo deveria ser caracterizado segundo suas necessidades de graus-dia, mas isto nem sempre ocorre. Em muitos casos, eles recebem classificação de precocidade na própria origem (superprecoce, precoce, normal, etc.) sem a devida comprovação experimental.
Além disso, em geral, há grande demanda por híbridos mais precoces, para semeaduras antecipadas no final do inverno, em safrinha de fim de verão-outono ou para expansão de áreas em regiões de altitude. No sul do Brasil, o cultivo em safrinha teve incremento nas últimas décadas, em função da antecipação de cultivo na primavera ou para lavouras de milho em sucessão ao feijão de primavera ou fumo.
Visando estabelecer critérios para caracterizar genótipos de milho segundo grupos de precocidade, Fancelli e Dourado Neto (1997) citados por Gadioli et al. (2000) descreveram que híbridos tardios necessitam acima de 890 graus-dia para o florescimento, precoces necessitam mais de 831 e menos de 890 graus-dia, enquanto superprecoces necessitam menos de 830 graus-dia. Este procedimento é necessário, mas sua implantação em escala operacional necessita critérios científicos claros, além de suporte em pesquisas de campo.
Como a temperatura atua diretamente no metabolismo básico das plantas seu efeito se reflete, amplamente, sobre a evolução de uma cultura no campo. Assim, o acúmulo de graus-dia também pode ser empregado como parâmetro para estimar o crescimento do índice de área foliar (IAF), como demonstra a Figura 7.
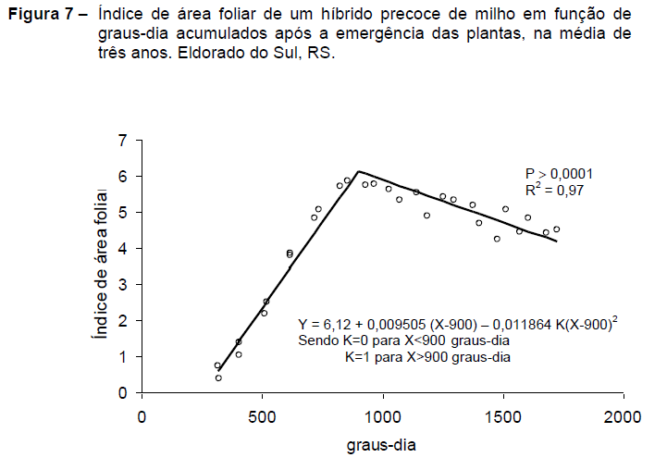
Baseado no modelo segmentado de Brisson et al. (1990), Müller et al. (2005) observaram que o crescimento do IAF do milho é linear entre o estádio de quatro folhas expandidas e o final da expansão da folha bandeira, que corresponde à antese.
No Rio Grande do Sul, um híbrido classificado como precoce necessitou aproximadamente 900 graus-dia (acima de 8ºC) para atingir o máximo IAF, a partir da emergência das plantas. A partir do IAF máximo inicia-se uma lenta senescência de folhas até próximo à maturação fisiológica, com cerca de 1800 graus-dia após a emergência.
A possibilidade de estabelecer modelos de crescimento do IAF a partir de uma única variável, de fácil obtenção como é a temperatura do ar, permite cumprir o primeiro passo importante da modelagem do crescimento e da formação do rendimento do milho.
Uma vez calculado o IAF, é possível estimar a produção de biomassa da cultura em função da radiação fotossinteticamente ativa interceptada pelo dossel e, com o índice de colheita, projetar o rendimento potencial de grãos (FRANÇA, 1997; MÜLLER; BERGAMASCHI, 2005).
Funções lineares são de fácil aplicação, sobretudo ao serem inseridas em modelos complexos, como são os modelos analíticos ou determinísticos. A função principal destes é a simulação dos principais processos que ocorrem ao longo do ciclo das culturas, e que resultam na produção de biomassa, de grãos ou frutos.
Entretanto, em geral, os processos biológicos não seguem funções lineares. Suas respostas às condições do ambiente e a própria evolução dos fenômenos no tempo são, tipicamente, não lineares. Assim sendo, em geral, os modelos lineares representam simplificações dos processos biológicos.
E isto se traduz em falhas na interpretação e nas aplicações dos princípios que regem as relações clima-planta, como o próprio uso de graus-dia para a simulação do desenvolvimento das plantas. (BONHOMME, 2000).
A Figura 8 representa a evolução do índice de área foliar de híbridos precoces de milho no Rio Grande do Sul, segundo Bergamaschi et al. (2006).
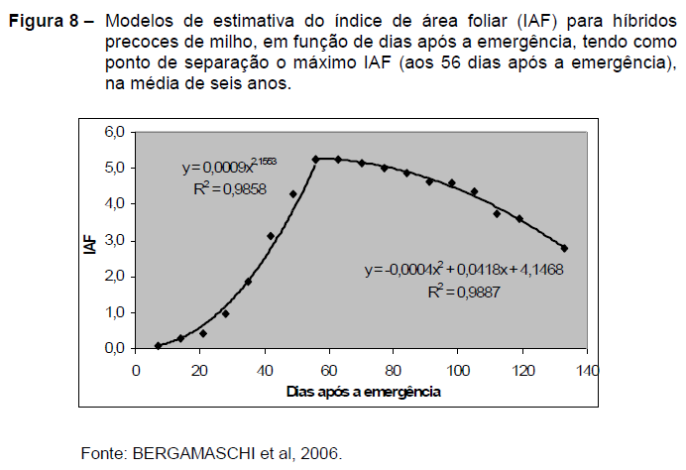
A base de dados é aquela utilizada na Figura 7, porém com maior número de anos. O ajuste de um segmento exponencial para o crescimento vegetativo e um segmento quadrático a partir do máximo IAF permitiu abranger todo o ciclo das plantas, da emergência à maturação fisiológica. A ampliação de três para seis anos nos dados de origem também contribuiu para que houvesse alta precisão nos ajustes.
Por outro lado, o uso de número de dias do calendário como variável independente, ao invés do critério de tempo térmico, restringe a validade do modelo a ambientes com climas semelhantes àqueles em que os dados foram gerados. Embora tenham sido utilizados dados de seis anos, haverá discrepâncias de estimativas sempre que o modelo for utilizado em locais ou épocas com condições térmicas diferentes.
Mesmo assim, o modelo não linear apresentado na Figura 3 se mostra coerente e lógico para simular a evolução do IAF no crescimento (até o florescimento) e seu lento declínio pela senescência gradual das folhas no período reprodutivo.
O emprego de graus-dia acumulados após a emergência, ao invés de dias do calendário, poderia tornar o modelo de uso mais amplo, para híbridos e arranjos de plantas semelhantes àqueles utilizados por Bergamaschi et al. (2006).
Se você tem interesse em saber mais sobre a Cultura do Milho, te convido a conhecer a plataforma da AgricOnline. Ao fazer a sua assinatura, você tem acesso ilimitado a todos os cursos da plataforma. São cursos que vão desde produção vegetal, produção animal, mercado e carreira.
Ao término de cada curso, você tem direito ao certificado com a carga horária de cada curso, clique no link para conhecer.

Ou clique no link:
https://go.agriconline.com.br/pass/?sck=portal
Fonte
BERGAMASCHI, Homero; MATZENAUER, Ronaldo. O Milho e o Clima. 1ª ed. Porto Alegre - RS: Emater/RS-Ascar, 2014.



