Influência da Nutrição no Aumento do Teor de Sólidos no Leite


Introdução
Certamente a comercialização do leite é a principal fonte de renda da maioria das propriedades leiteiras, sendo de suma importância o volume de leite produzido. Porém, assim como já ocorreu em vários países de pecuária leiteira mais desenvolvida, existe uma forte tendência pela remuneração por composição e qualidade sanitária do leite e, não apenas por volume de leite produzido.
Mesmo não sendo ainda uma realidade nacional, a remuneração com bonificações e penalizações pelos teores de sólidos no leite (teores de gordura e proteína) e sanidade (Contagem Bacteriana Total - CBT e Contagem de Células Somáticas - CCS) já é uma realidade que ocorre, por exemplo, em vários laticínios e cooperativas progressistas nas regiões Sul e Sudeste do Brasil.
Essas novas formas de remuneração seguem as demandas e exigências de mercado. E o que se tem observado em vários países, inclusive no Brasil, é um maior interesse dos consumidores em adquirir produtos ditos mais saudáveis. No que diz respeito à cadeia de lácteos, vêm ocorrendo uma crescente busca por produtos com menores teores de gordura e maiores teores de proteína. Consumidores de países desenvolvidos estão comprando muito mais queijos, sorvetes, iogurtes e leite desnatado ou com baixo teor de gordura do que anos atrás.
Essas necessidades e demandas de mercados refletem sobre toda cadeia de lácteos, podendo acarretar alterações na seleção das características produtivas. Por exemplo, particularmente na Europa e na América do Norte tem se observado maior ênfase na seleção para proteína no leite devido ao consumo crescente de queijos.

Genética, componentes e volume de leite
Nutrição, manejo e genética são os principais meios para se alterar a composição do leite. Mudanças na composição do leite através da genética são obtidas a longo prazo, principalmente pelo longo intervalo de gerações na espécie bovina, se comparadas às alterações produzidas por práticas de nutrição e de manejo, que geralmente ocorrem de forma quase imediata. Por outro lado, as mudanças obtidas pela genética são permanentes, ao contrário das alterações proporcionadas pelas práticas de nutrição, que são transitórias.
Entre os componentes do leite, a gordura apresenta a maior variabilidade, com mediana variabilidade para a proteína e menor para a lactose. Segundo dezenas de publicações científicas, as produções de gordura e de proteína apresentam medianas herdabilidades (0,25 a 0,30), enquanto que as porcentagens de gordura e de proteína apresentam altas herdabilidades (0,45 a 0,50). Mas apesar das altas herdabilidades, selecionar para altas porcentagens de gordura e de proteína pode causar problemas, em função das porcentagens serem negativamente correlacionadas com o volume de leite.
Na Tabela 1 estão descritas as mudanças esperadas na composição do leite por diferentes critérios de seleção.
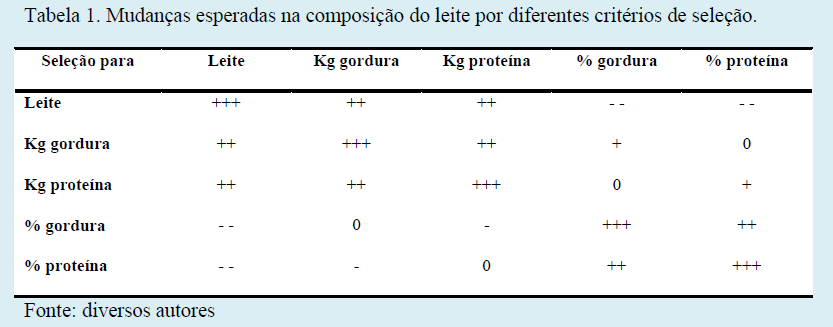
Na tabela anterior fica evidenciado que selecionar para quilogramas de sólidos (gordura e/ou proteína) parece ser a melhor estratégia, pois além de maximizar a resposta em quilogramas de gordura e proteína, o volume de leite também é incrementado. Por outro lado, se optássemos por selecionar para teores de gordura e/ou proteína, mesmo com respostas máximas em teores de sólidos, teríamos como resposta correlacionada uma diminuição no volume de leite.
Portanto as três características produtivas (leite, gordura e proteína) deveriam ser consideradas em conjunto na seleção, preferencialmente num índice de seleção, incluindo a variabilidade e o peso econômico desejado para cada característica.
Composição do leite e diferenças entre raças
A composição normal do leite bovino contém cerca de 12,5% de sólidos, como demonstrado na Tabela 2.

A composição do leite considerada normal para vacas leiteiras pode variar em função da raça dos animais. A raça Holandesa apresenta menores percentuais de sólidos, mas maiores produções totais de gordura e proteína. Já as raças Jersey e Pardo-Suíço, apesar dos altos percentuais de gordura e proteína, apresentam produções totais destes componentes ligeiramente inferiores aos encontrados na raça Holandesa.
Demonstrando as afirmações acima, nas Tabelas 3 e 4 estão relacionadas as produções de leite, de gordura e de proteína, bem como as porcentagens de gordura e de proteína, tanto em rebanhos norte-americanos, como em rebanhos canadenses, ambos em controle leiteiro. Em rebanhos leiteiros paranaenses, segundo dados de Almeida et al. (1999) e Ribas et al. (1999), estas mesmas tendências são observadas.
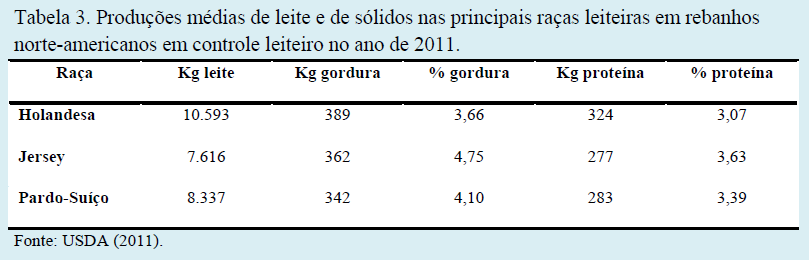
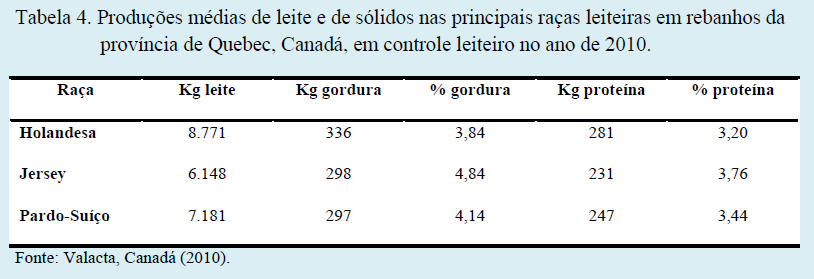
Além de características raciais e genéticas, as técnicas de manejo, condições climáticas e principalmente a nutrição podem alterar a composição do leite. Por se tratar de um fator econômico oneroso para produção de leite e causar grandes impactos sob o volume de leite produzido e seus teores proteicos e lipídicos, a nutrição de bovinos leiteiros é alvo de discussões já a longa data por pesquisadores, técnicos e produtores.
Depressão da gordura do leite
Existem dois grupos principais de dietas que podem causar a depressão da gordura no leite (DGL). O primeiro grupo envolve dietas que fornecem grandes quantidades de carboidratos prontamente digestíveis e reduzidas quantidades de componentes fibrosos, tais como dietas com alta proporção de grãos e baixa proporção de forragem. Dietas onde o conteúdo de fibra é adequado, mas esta fonte de fibra é peletizada ou demasiadamente picada também são incluídas nesta categoria, já que estes processos reduzem a capacidade da fibra de manter a atividade normal do rúmen (fibra efetiva).
O segundo grupo de dietas que induzem a DGL são suplementos dietéticos contendo óleos polinsaturados, tais como óleos de origem vegetal e de origem marinha. Como a suplementação de óleos de origem marinha é restrita pela proibição imposta pelo MAPA por conta da encefalite espongiforme bovina (BSE), nossa preocupação deve ser direcionada aos óleos de origem vegetal e em particular às gorduras com altas proporções do ácido graxo linoléico (C18:2), tais como o óleo de soja e produtos derivados.
Em ruminantes os lipídios presentes na dieta são extensivamente alterados pelos microrganismos do rúmen (Jenkins, 1993). Esta alteração é uma consequência da biohidrogenação dos ácidos graxos polinsaturados presentes na dieta, um processo que tende a tornar a gordura do leite mais saturada, mas que também resulta na formação e secreção de inúmeros ácidos graxos do tipo "trans" no leite, com destaque aos isômeros de CLA (ácido linoléico conjugado) cis-9 trans-11 e trans-10 cis-12.
Devido a estas e outras transformações que ocorrem no rúmen, mais de 400 tipos de ácidos graxos já foram identificados na gordura do leite de bovinos. Os ácidos graxos presentes em maior concentração são o ácido palmítico (C16:0) e o ácido oléico (C18:1 cis-9), sendo que a soma dos dois representa cerca de 50% do total.
Os ácidos graxos secretados no leite podem ter duas origens: síntese de novo nas células epiteliais mamárias ou circulação sanguínea. Ácidos graxos de cadeia curta (4-8 carbonos) e média (10-16 carbonos) são sintetizados quase que exclusivamente pela síntese de novo na glândula mamária, predominantemente a partir do acetato (produto da degradação de carboidratos no rúmen). Em contrapartida, os ácidos graxos de cadeia longa, com 18 ou mais carbonos, são oriundos exclusivamente da circulação.
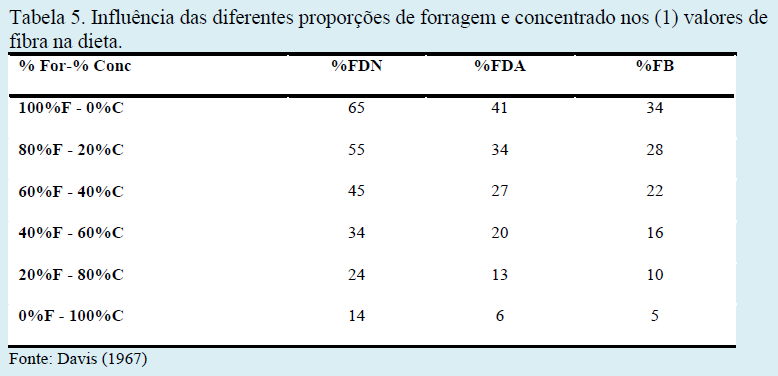
Características físicas dos alimentos volumosos, a relação entre volumoso e concentrado e, frequência na alimentação afetam diretamente o ambiente ruminal, que por sua vez reflete sobre a produção animal, podendo alterar não somente o perfil de ácidos graxos do leite, mas também promover uma redução do teor (%) e da secreção (kg/dia) de gordura do leite.
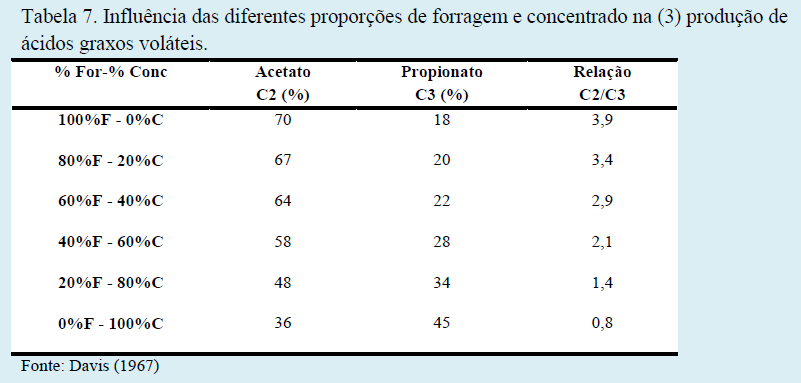
Os teores adequados de fibra na dieta de bovinos são primordiais para um adequado desempenho ruminal e manutenção de seu pH. Diferentes proporções entre forragem e concentrado na dieta acarretam distintos teores de fibra em detergente neutro (FDN), fibra em detergente ácido (FDA) e fibra bruta (FB), em respostas ruminais distintas e em produções de ácidos graxos de cadeia curta em proporções variadas (Tabelas 5, 6 e 7).
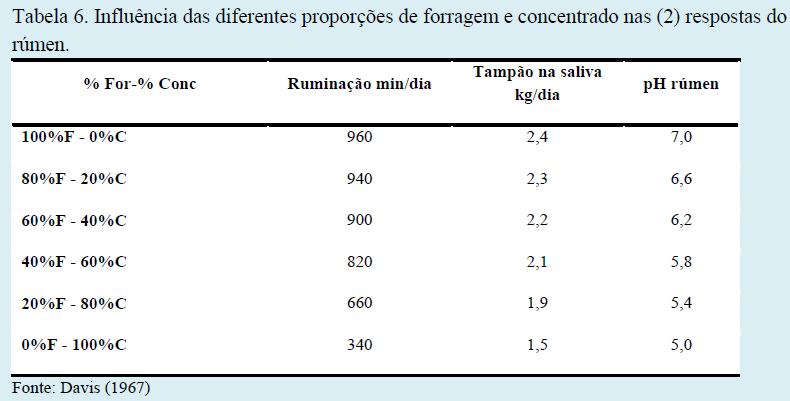
Bauman & Griinari (2003) avaliaram o fornecimento de dietas contendo baixa fibra e seus efeitos sobre a produção de leite, gordura e ácidos graxos voláteis comparados a um grupo controle. Os autores demonstraram que houve redução na produção (g/dia) e no teor (%) de gordura no leite, mas que a produção diária de acetato (importante precursor para produção da gordura do leite) não foi alterada, ao contrário do propionato que foi produzido em quantidades bem acima dos níveis encontrados na dieta controle (13,3 vs. 31,0 moles/dia), como demonstrado na tabela seguinte.
O segundo grupo de dietas que induzem a DGL são suplementos dietéticos contendo óleos polinsaturados, tais como óleos de origem vegetal e de origem marinha. Óleo de peixe e outros óleos de origem marinha são, em geral, caracterizados pela presença de quantidades significativas de dois ácidos graxos polinsaturados: o eicosapentaenóico (C20:5) e o docosahexaenóico (C22:6). Ao contrário dos óleos vegetais, os óleos de origem marinha irão induzir a DGL mesmo quando as dietas contêm níveis adequados de fibra.
Várias teorias têm sido propostas para explicar a DGL induzida pela dieta, e alterações nos processos microbianos do rúmen são à base de todas estas teorias. Uma das teorias advoga que alterações na fermentação ruminal resultam numa inadequada produção ruminal de acetato e butirato para dar suporte à síntese de novo da gordura do leite na glândula mamária.
De fato, há grandes variações percentuais de ácidos graxos voláteis do rúmen quando fornecemos dietas de baixa fibra (Tabela 7). Como demonstrado por Bauman & Griinari (2003) na Tabela 8, a concentração molar de acetato diminui, a de propionato aumenta e, por consequência, a relação acetato:propionato cai drasticamente. Entretanto, embora a proporção molar de acetato seja reduzida, a produção de acetato (moles/dia) não é afetada. Portanto, a redução na relação acetato:propionato é uma consequência da maior produção de propionato.
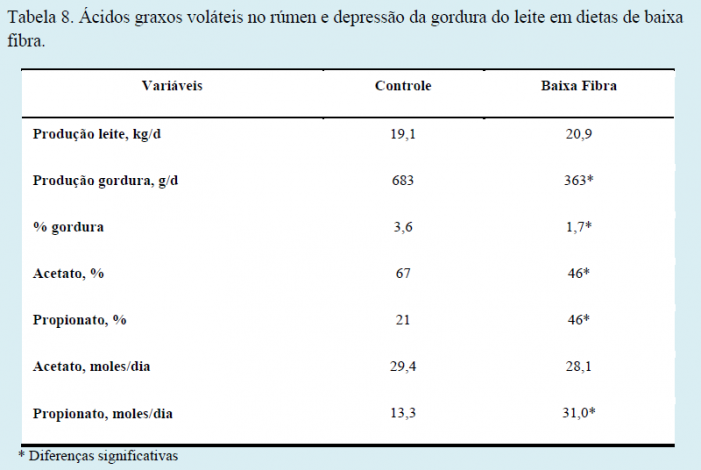
A possibilidade de que a disponibilidade de acetato poderia estar limitando a síntese de gordura do leite também foi examinada por experimentos onde acetato exógeno foi administrado a vacas recebendo dietas de baixa fibra. As respostas ao fornecimento de acetato foram modestas ou inexistentes em tais estudos. Em resumo, já que a produção de acetato não é reduzida em dietas com baixa fibra, esta teoria foi abolida, já que ela não poderia explicar a DGL.
A teoria mais aceita atualmente para explicar a DGL é conhecida como “Teoria da Biohidrogenação” ou "Teoria dos Ácidos Graxos Trans". Esta teoria postula que a DGL não é causada pela escassez de precursores lipogênicos para a glândula mamária. Ela afirma que a síntese mamária da gordura do leite é inibida diretamente por tipos específicos de ácidos graxos, produzidos a partir da biohidrogenação parcial dos lipídios da dieta sob certas condições ruminais.
A queda no pH ruminal (proporcionado pelo fornecimento de dietas de baixa fibra ou de baixa efetividade física) altera as rotas de biohidrogenação ruminal (Figura 1), produzindo um ácido graxo específico, CLA trans-10 cis-12, cuja concentração aumenta significativamente no leite de vacas com DGL (Tabela 9). Estudos posteriores, onde este isômero de CLA foi sintetizado industrialmente e infundido no abomaso de vacas em lactação, comprovaram sua grande capacidade de reduzir a secreção de gordura do leite.

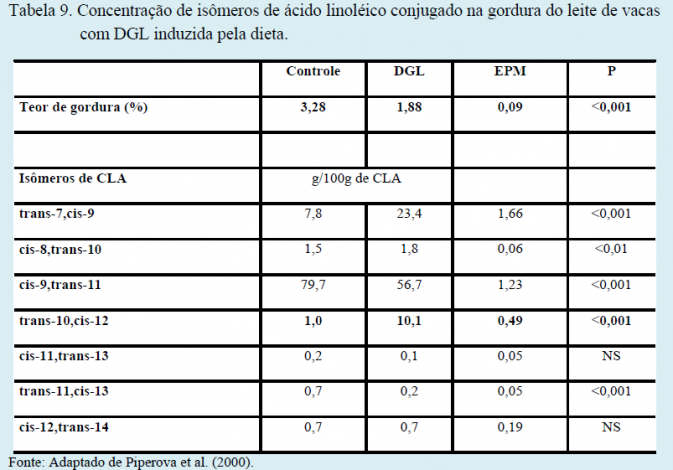
De grande relevância, estudos recentes demonstraram que a formação do CLA trans-10 cis-12 no rúmen só ocorre quando duas condições estão presentes: baixo pH ruminal (ex.: dietas de baixa fibra) e presença de lipídios polinsaturados na dieta (ex.: dietas suplementadas com grãos de oleaginosas, óleo de milho, sais cálcicos de ácidos graxos a partir do óleo de soja, etc.). A ausência de qualquer uma das condições não resultará em DGL.
Muitas pesquisas têm enfocado o aumento na quantidade de ácido linoléico conjugado (CLA) e de ácido transvaccênico (trans-11 C18:1) na gordura do leite pelos benefícios relatados na saúde humana. Segundo Whitlock et al. (2002) certos isômeros de CLA, especialmente o cis-9, trans-11, têm sido identificados como tendo propriedades anticarcinogênicas, além de antiaterosclerose, antitrombótico, hipocolesterolêmico, previne o diabetes e apresenta efeito imunoestimulatório. Outros isômeros de CLA, incluindo o trans-10, cis-12, parecem ainda possuir propriedades antiobesidade.
Monitoramento do FDN e do FDNfe da dieta
Respeitar as recomendações e exigências de FDN e FDNfe (fibra em detergente neutro fisicamente efetiva) de vacas leiteiras em suas diversas fases produtivas, são primordiais para a manutenção do pH ruminal, favorecendo assim a fermentação microbiana e a biohidrogenação adequado de ácidos graxos polinsaturados, ou seja, sem a formação de CLA trans-10, cis-12, que está diretamente relacionado a DGL.
As recomendações do penúltimo NRC para bovinos leiteiros (NRC, 1989) sugeriam que a dieta de bovinos leiteiros deveria conter no mínimo 28% de FDN e 21% de FDA. Para vacas de alta produção a recomendação era de no mínimo 25% de FDN e 19% de FDA, e que 75% da FDN total da dieta deveria ser proveniente de forragem.
Por conta do uso crescente de subprodutos fibrosos nas dietas de vacas leiteiras, Mertens (1997) e o NRC (2001) estabeleceram que o valor efetivo médio do FDN não forragem é de 50% do FDN forragem. Desta forma, as recomendações seguem a seguinte regra: para cada redução de 1% no FDN forragem abaixo de 19%, a concentração do FDN total da dieta deve aumentar em 2%, enquanto que a %CNF (carboidratos não fibrosos) deve diminuir em 2% (Tabela 10).
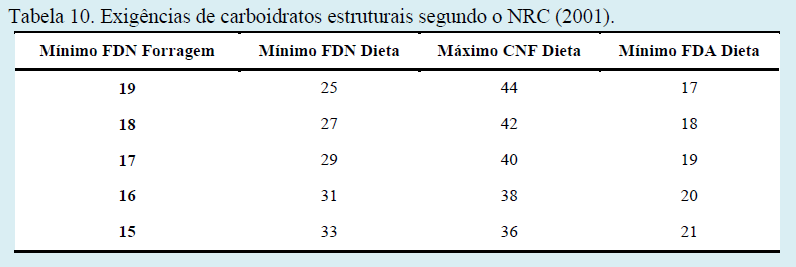
Segundo as recomendações do NRC (2001), os valores para FDN, FDA e CNF sugeridos são válidos quando três condições específicas são observadas: uso de dieta total misturada (TMR), tamanho de partícula adequado e que o milho moído seja a fonte predominante de amido utilizado na dieta. Em outras palavras, o NRC (2001) sugere que a concentração de FDN na dieta deve ser mais alto quando se usa forragem excessivamente picada ou processada e, em dietas que contenham altas proporções de subprodutos ricos em FDN não forragem, tais como casquinha de soja, refinasil, polpa cítrica, etc.
As recomendações de FDN podem sofrer ajustes em função de aspectos qualitativos da dieta, tais como fontes de amido disponíveis, tamanho de partícula da forragem, fibra efetiva da dieta, suplementação de tampões e do manejo alimentar adotado.
Em se tratando de diversas fontes de amido, as exigências de FDN aumentam quando a disponibilidade do amido no rúmen também aumenta. Essa maior ou menor disponibilidade de amido no rúmen é relevante quando se compara alguns alimentos que são fonte desse nutriente, como por exemplo: grãos moídos vs. grãos quebrados, grãos floculados vs. Grãos secos, grãos de alta umidade vs. grãos secos, grãos de cereais de inverno vs. milho, etc.
O conceito de fibra em detergente neutro fisicamente efetiva (FDNfe) surgiu para atender a exigência mínima de fibra que mantém a porcentagem de gordura no leite em níveis adequados. No ambiente ruminal sabe-se que nem todo FDN presente na dieta tem atividade efetiva no rúmen e no metabolismo animal, ou seja, podem não apresentar características físicas desejadas (tamanho e densidade de partículas), estímulo a atividade de mastigação e produção de saliva, manutenção de pH e neutralização de ácidos produzidos no rúmen.
Mertens (1997) propôs um método para calcular o FDNfe, o qual considera a %FDN na dieta e a quantidade de partículas retidas em uma peneira maior que 1,18 mm (FDNfe = %FDN x fef). O fator de efetividade física varia de 0 a 1 (Tabela 11).
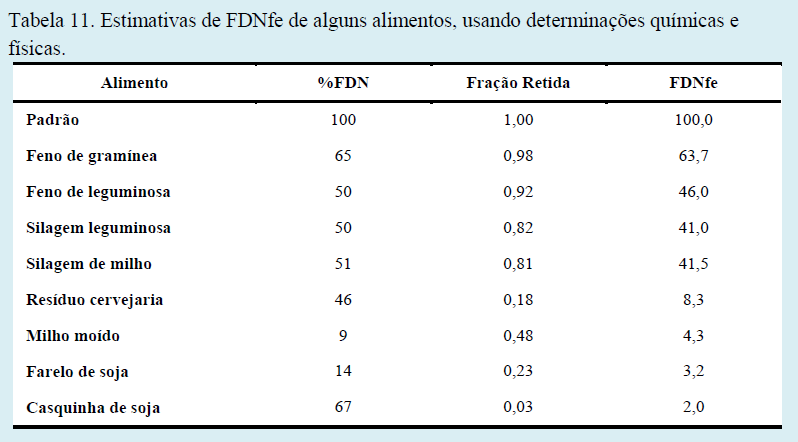
Não é incomum a existência de alimentos com quantidades de FDN muito semelhantes, mas que apresentem diferentes valores de efetividade física.
As exigências de FDNfe para bovinos leiteiros não foram definidas pelo NRC (2001). Já Mertens (1997) estimou que a exigência de FDNfe para vacas leiteiras é de 22%MS para manter um pH ruminal de 6,0 e de 20%MS para manter o teor de gordura no leite em 3,40%. Na nossa modesta experiência ao formular dietas em rebanhos leiteiros paranaenses, tentamos respeitar o limite de 21% de FDNfe para dietas de vacas leiteiras de alta produção, desde que os níveis de amido não sejam excessivos (até 26%MS).
Avaliar o tamanho de partícula na dieta é primordial para garantir a efetividade da fibra e evitar que os animais selecionem alimentos mais concentrados. Vacas têm capacidade de selecionar partículas de forragem maiores que 5 cm de comprimento. Assim, se estiver ocorrendo seleção dos alimentos, é recomendável que a forragem seja picada entre 2,5 e 5 cm.
O quadro de subacidose é caracterizado por pH ruminal abaixo de 5,8 e uma das prováveis causas é a ingestão excessiva de carboidratos de rápida fermentação, decorrente de desbalanceamento da dieta ou seleção do alimento por parte do animal. Alguns sinais que podem facilitar a identificação de acidose ruminal: consumos de MS e produções de leite variando muito de um dia para outro, aspecto do esterco inconsistente, falta de ruminação, consumo voluntário de bicarbonato, acidose sistêmica provocando laminite e depressão no aspecto geral das vacas, além de obviamente queda nos teores de gordura do leite.
Outra forma de diagnosticar a ocorrência da DGL em um rebanho, é monitorar a relação gordura:proteína do leite. Como já relatado anteriormente os teores de gordura no leite apresentam maiores variações que os teores de proteína, além de ser um componente que permite maior manipulação, principalmente através da dieta. Portanto quando se diz que houve uma inversão entre a relação %gordura:%proteína, o que provavelmente ocorreu foi uma redução nos teores de gordura no leite e não um aumento nos teores de proteína.
A relação considerada ideal para a relação %gordura/%proteína na raça Holandesa é de 1,10, mas valores entre 1,00 e 1,20 são aceitáveis. Se num rebanho mais de 10% das vacas em lactação apresentarem 0,2% de gordura abaixo do teor de proteína (exemplo; 2,8%G e 3,0%P) isso pode ser um indício de acidose no rebanho e/ou desbalanceamento da dieta.
Impacto do aumento dos níveis de PB da dieta e nitrogênio ureico no leite
A proteína é um nutriente essencial para produção de leite e que influencia a ingestão de alimentos pelo animal. No leite de vacas, a proteína verdadeira constitui cerca de 94% do nitrogênio total e o restante é representado pelo nitrogênio não proteico (NNP).
O monitoramento dos teores de nitrogênio ureico no leite (NUL) é uma ferramenta que tem se tornado cada vez mais utilizada, por estar diretamente relacionada ao teor proteico e energético da dieta e a excreção de N pelo animal. Tem-se buscado valores de NUL que representem um ponto de equilíbrio entre ingestão de proteína e produção de leite sem grandes perdas de N, ou seja, maior eficiência na utilização de N (EUN).
Na última década, a maioria das publicações norte-americanas passou a sugerir metas para NUL valores entre 8 e 12 mg/dL (Kohn et al., 2002; Rajala-Schultz & Saville, 2003), com algumas publicações mais recentes sugerindo valores ainda menores; entre 7 a 10 mg/dL.
No Brasil minha sugestão é mais conservadora; valores entre 10 e 14 mg/dL parecem ser ainda os mais indicados (Almeida, 2012). Embora vários rebanhos norte-americanos estejam tendo sucesso em conciliar baixos níveis dietéticos de PB (inferiores a 16,0%PB) e baixos valores de NUL (inferiores a 10 mg/dL) com altas produções de leite, acreditamos que no Brasil a redução muito exagerada dos teores de PB das dietas poderão comprometer a produção de leite. Esta distinção ocorre pela impossibilidade de uso de suplementos proteicos de origem animal (naturalmente ricos em PNDR), pela não-disponibilidade de suplementos proteicos de origem vegetal com baixa degradabilidade ruminal, bem como pela não-popularidade na suplementação de aminoácidos sintéticos (metionina e lisina) em dietas brasileiras.
O monitoramento mensal do NUL pode ser uma importante ferramenta no manejo de rebanhos leiteiros, porque (1) o excesso no consumo de proteína (N) pode comprometer a eficiência reprodutiva; (2) suplementos proteicos são ingredientes caros; e (3) excessos na excreção de N têm um impacto ambiental negativo (Jonker et al., 1998).
Entre as práticas nutricionais que podem favorecer o aumento do NUL, podemos destacar: redução exagerada dos níveis dietéticos de amido pela inclusão de fontes de carboidratos não amiláceos, como casca de soja e polpa cítrica; o uso de silagem de milho “nova” (menos de 3 meses de ensilagem); pastagens novas e muito adubadas, principalmente de inverno; pré-secado úmido e com alto %PB; fornecimento do grão de milho mais grosseiro (quebrado ao invés de moído).
Impacto do estresse calórico na composição do leite
Animais da raça Holandesa são menos tolerantes a temperatura mais elevada que outras raças leiteiras européias, como Jersey e Pardo-Suíço, e muito menos tolerantes que grupamentos raciais com sangue zebuíno, como Girolando e Gir Leiteiro. Vacas Holandesas submetidas a estresse calórico reduzem sua produtividade de forma expressiva. Num levantamento preliminar conduzido pelo nosso grupo de pesquisa (dados não publicados), realizado em 26 rebanhos paranaenses em controle leiteiro oficial na região de Arapoti, Paraná, a diferença entre o mês de maior produtividade média (setembro) e o mês de menor produtividade média (fevereiro) alcançou 8 litros diários!
O estresse calórico impacta negativamente vários parâmetros na atividade leiteira, tais como produção de leite, qualidade e composição do leite, saúde ruminal, crescimento e reprodução. St. Pierre et al. (2003) estimaram as perdas da indústria leiteira norte-americana devido ao estresse calórico em 900 milhões de dólares anuais! Avanços no manejo com a adoção de práticas de resfriamento e estratégias nutricionais podem amenizar alguns dos impactos negativos do estresse calórico em bovinos leiteiros, mas as produções continuam a declinar nos meses mais quentes.
O mecanismo biológico pelo qual o estresse calórico impacta a produção e a reprodução é parcialmente explicado pela depressão no consumo alimentar, mas também inclui outros fatores tais como mudança no status endócrino, reduções na ruminação e na absorção de nutrientes e aumento nas exigências de mantença. As reduções tanto nos teores de gordura como nos de proteína verdadeira relatadas nas épocas mais quentes do ano pela indústria leiteira norte-americana se situa entre 0,2 e 0,3%. Infelizmente desconhecemos a existência de levantamentos similares pela indústria leiteira nacional.
De acordo com Nayeri et al. (2011) existem algumas estratégias que podem ser adotadas para minimizar o impacto do estresse calórico na produção e composição do leite. Uma das mais tradicionais é aumentar a densidade energética da dieta, seja pela redução de fibra, pelo aumento do concentrado ou ainda pela suplementação de gorduras. Estas práticas devem ser adotadas com cuidado, pois se aumenta a predisposição das vacas em estresse calórico à acidose ruminal. A manipulação da Diferença Catiônica-Aniônica da Dieta (DCAD), buscando valores francamente positivos durante a lactação (+20 a +30 meq/100g MS), bem como o aumento nos níveis dietéticos do mineral potássio no verão (1,4 a 1,6%MS) também têm sido sugeridos.
Outras práticas nutricionais e de manejo comumente recomendadas são o fornecimento preferencial de alimento (60-70% da dieta) nas horas mais frescas do dia, a limpeza mais frequente dos cochos com maior número de tratos diários, o fornecimento de concentrado mais parcelado ao longo do dia e a inclusão de 2 a 2,5 kg de caroço de algodão.
Impacto de alguns aditivos nutricionais na composição do leite
Monensina
A monensina, assim como outros ionóforos, é um antibiótico, produzido por fungos do gênero Streptomyces cinnamonensis, com reconhecida eficácia nas explorações pecuárias. Os dados de literatura são abundantes e extremamente consistentes para o efeito dos ionóforos, tanto em bovinos de corte como em bovinos leiteiros. A maior parte dos experimentos foi conduzida com monensina sódica.
O efeito mais notório dos ionóforos é o aumento da retenção de energia fermentada no rúmen devido a uma alteração no padrão de fermentação, com maior produção de propionato (C3) em relação a acetato (C2) e decorrente diminuição das perdas através do metano. Os ionóforos também parecem diminuir a degradação da proteína ruminal e, portanto, reduzindo a degradação de peptídeos e aminoácidos e resultando em menor produção de amônia. Por último, os ionóforos (particularmente a monensina) diminuem a ocorrência de distúrbios metabólicos, como acidose e timpanismo, pela menor concentração de ácido lático e menor produção de mucopolissacarídeos que dão estabilidade à espuma.
Os efeitos nas produções de gordura e de proteína são heterogêneos e dependem de fatores dietéticos. Mas na meta-análise que reuniu 36 artigos ou 77 experimentos envolvendo monensina e dados de produção (Duffield et al., 2008), a suplementação com monensina aumentou a produção de proteína, diminuiu as porcentagens de gordura e de proteína no leite, e não apresentou efeito na produção de gordura.
Durante muito tempo atribuiu-se à menor produção de acetato no rúmen o efeito de redução na gordura no leite, já que de fato o acetato é o principal precursor da gordura do leite. Mas hoje se sabe que na realidade o efeito depressor da monensina na gordura do leite é explicado pela diminuição das taxas de biohidrogenação dos ácidos graxos insaturados no rúmen, o que contribui no aumento das concentrações do ácido linoléico conjugado trans-10 cis-12, potente inibidor da síntese de gordura do leite na glândula mamária (Bauman & Griinari, 2003). Duffield et al. (2008) confirmaram que a suplementação com monensina aumenta a proporção de ácido linoléico conjugado (CLA) no leite. Maiores consumos de fontes lipídicas insaturadas na dieta e altas inclusões de monensina (ao redor de 400 ppm) exacerbam a diminuição da gordura do leite com o tratamento com monensina.
Dosagens recomendadas de monensina para vacas em lactação estão entre 11-22 mg/kg de MS consumida, ou 250-400 mg/vaca/dia (Hutjens, 2010). No Brasil, devido ao impacto negativo da monensina na gordura do leite e ao potencial decréscimo no consumo de ração (R.P. Nogueira, comunicação pessoal), dosagens mais próximas do limite inferior são sugeridas (11-15 mg/kg).
Leveduras
Leveduras vêm sendo fornecidas aos animais há mais de cem anos. Embora existam aproximadamente 500 diferentes espécies de leveduras, a mais comum na suplementação dos bovinos é a Saccharomyces cerevisiae.
Segundo Santos & Greco (2012), de modo geral, o efeito das leveduras vivas e cultura de leveduras se dá por alterações no ambiente ruminal, com mudanças na população microbiana que favorecem o crescimento de microrganismos celulolíticos e daqueles utilizadores do ácido lático.
De maneira geral, a adição de leveduras a dieta de bovinos leiteiros resulta em aumento na ingestão de MS, estabilidade do pH ruminal (utilização do ácido lático), melhora a digestão de fibra no rúmen, aumenta a produção de leite (entre 1 a 1,2 kg/dia), aumenta a concentração de gordura no leite e produção de componentes do leite (Santos & Greco, 2012).
De acordo com os autores citados anteriormente, a dose típica recomendada para bovinos leiteiros é de 1 a 2 x 1010 unidades formadoras de colônias por dia para bovinos adultos. A conversão desses valores em g/dia/animal depende da concentração de leveduras presentes no produto comercial, mas os valores podem variar de 5 a 120 g/dia.
Aminoácidos Metionina e Lisina
A alimentação de vacas leiteiras especializadas baseia-se na oferta dos nutrientes exigidos pelo animal de forma a atender suas necessidades adequadamente, sem que ocorra a falta ou o excesso de algum nutriente. A proteína, quando em excesso na dieta, além de promover a maior produção de fezes, também é responsável pela ineficiência de utilização do nitrogênio para a produção de leite (Jonker et al., 2002).
Com isso, torna-se imprescindível a utilização de dietas adequadamente formuladas, principalmente quanto aos teores de proteína e seus aminoácidos limitantes (metionina e lisina), que atendam às necessidades da vaca leiteira especializada, resultando potencialmente em maior quantidade e qualidade de leite, redução dos impactos negativos ao ambiente e diminuição dos custos com a alimentação (NRC, 2001).
Robinson (2010) numa revisão de literatura que reuniu 54 experimentos comparando a suplementação de lisina e/ou metionina com dietas controle, concluiu que a manipulação das proporções de lisina e metionina da proteína duodenal pela suplementação de metionina protegida, ou lisina combinada a metionina, apresentou efeitos positivos, mas modestos, na melhoria do desempenho produtivo, bem como na redução do impacto ambiental de vacas leiteiras. Já a suplementação isolada de lisina sintética tem impacto até negativo.
Robinson (2010) justificou os resultados modestos argumentando que a contribuição da proteína microbiana na proteína metabolizável é tão grande, e que o perfil de aminoácidos desta proteína já é tão similar com o perfil de aminoácidos das proteínas do leite, que mesmo suplementando um aminoácido específico, o benefício gerado em resposta animal é geralmente pequeno. Ainda assim, na meta-análise conduzida, o autor concluiu que a suplementação com metionina aumentou em 1,3% a secreção de energia no leite, bem como os teores de gordura (+1,1%) e de proteína (+2,3%) no leite, além de aumentar em 2,1% a proporção de N dietético capturado como N no leite (eficiência de utilização do nitrogênio).
Nosso grupo de pesquisa conduziu recentemente dois experimentos avaliando a suplementação de metionina análoga (MFP®, Novus International Inc., EUA). No primeiro ensaio (Almeida et al., 2010) foram utilizadas 48 vacas Holandesas pareadas e o tratamento consistiu na suplementação de 20 g diárias de metionina análoga, por 4 semanas num delineamento de reversão simples.
A suplementação com metionina não alterou a produção de leite e seus componentes, mas observou-se uma tendência de maior teor de gordura nas vacas tratadas; 3,48% vs. 3,39%. Vacas suplementadas com metionina produziram leite com menores concentrações de nitrogênio ureico do que nas vacas controle; 16,83 e 17,44 mg/dL, respectivamente.
Num segundo experimento (Alegransi et al., 2011), 88 primíparas das raças Holandesa e Pardo-Suíça foram pareadas e suplementadas com 25 g diárias de metionina análoga, por 6 semanas, novamente num delineamento de reversão simples. Com a inclusão dietética de MFP®, a relação estimada entre lisina:metionina atingiu o valor desejável de 3,1:1 (6,40%Lis e 2,07%Met).
Não se observou diferença significativa para produção de leite entre vacas tratadas e não tratadas com metionina análoga; 34,2 vs. 33,3 kg/dia, respectivamente. Mas animais suplementados com MFP® produziram 53 g mais gordura do que vacas do grupo controle. Também se observou uma tendência de que vacas tratadas com metionina análoga produziram mais leite corrigido para 3,5% de gordura, 30 g mais proteína, e maiores teores de sólidos no leite.
Na nossa modesta opinião, há pelo menos três razões que limitam a inclusão de metionina análoga em dietas de rebanhos leiteiros brasileiros: a ainda restrita adoção de sistemas de pagamento de leite por qualidade, dados nacionais mostrando os benefícios da inclusão dietética de metionina ainda escassos e menor número de rebanhos de boa produtividade que justificam a suplementação.
Por outro lado, há no mínimo três razões que justificam a inclusão da metionina análoga em rebanhos nacionais: o fato do farelo de soja (pobre em metionina) ser o principal suplemento proteico em dietas de vacas leiteiras, a proibição (até exagerada) de suplementos proteicos de origem animal em dietas de ruminantes no Brasil e a não-disponibilidade no mercado brasileiro de alimentos proteicos de origem vegetal com baixa degradabilidade ruminal.
Por conta desta não disponibilidade de suplementos proteicos com maiores valores de proteína não-degradável no rúmen e da não popularidade da suplementação de aminoácidos protegidos (metionina em particular), há no Brasil uma cultura entre os nutricionistas de bovinos leiteiros que rebanhos ou lotes de vacas de alta produção deve ser suplementadas com altos teores de proteína bruta (ao redor de 17 a 18%PB). Não concordamos com esta excessiva margem de segurança, e acreditamos que sim é possível trabalhar com valores mais modestos (16 a 16,5%PB).
Bicarbonato de sódio e óxido de magnésio
Segundo o NRC (2001) compostos tais como bicarbonato de sódio e óxido de magnésio são incorporados às dietas em ordem de reduzir transtornos digestivos e manter o teor de gordura no leite, principalmente quando as dietas são ricas em grãos (carboidratos de alta fermentabilidade) e/ou pobres em ingredientes volumosos, fontes de fibra efetiva. Quando corretamente suplementados, estes tampões estabilizam o pH ruminal em torno de 6,2, permitindo um máximo crescimento das bactérias celulolíticas.
O óxido de magnésio, além de ser fonte do macromineral magnésio, é um alcalinizante (aumenta o pH ruminal) e parece atuar também na captura de metabólitos sanguíneos pela glândula mamária, aumentando o teor de gordura no leite produzido. A recomendação de inclusão do óxido de magnésio é de 1 parte deste alcalinizante para cada 2-3 partes de bicarbonato de sódio. Assim, quando da suplementação de óxido de magnésio, não se recomenda a exclusão do bicarbonato de sódio, mas sim a adição de ambos.
As recomendações de bicarbonato de sódio então entre 0,75-1,0% MS/dia, já o óxido de magnésio em 0,25% MS/dia. Segundo Almeida & Ostrensky (2011), em rebanhos confinados de mediana e alta produtividade, outra forma de suplementar bicarbonato de sódio às vacas é fornecê-lo ad libitum, em cochos exclusivos, além do bicarbonato já fornecido às dietas. Neste caso a função do bicarbonato ofertado nos cochos é de monitorar possíveis ocorrências de acidose ruminal subaguda. Como animais sem acidose normalmente não consomem voluntariamente o bicarbonato, quando isto acontece de maneira ávida por muitos animais, este fato pode chamar a atenção do produtor que a dieta está desequilibrada e que o nutricionista do rebanho precisa reformular as dietas.
Gordura protegida
Gordura protegida ou gordura inerte ruminal é um sal cálcico de um ácido graxo (AG), ou seja, um produto onde se combinam moléculas de AG com cálcio, para que o produto passe pelo rúmen sem sofrer dano extensivo pela ação da flora ruminal e sem interferir com esta. A ligação entre o cálcio e o AG não é quebrada por ação de enzimas microbianas e sim pelo pH do meio. Segundo Almeida & Ostrensky (2011) para a produção deste sal cálcico de AG podem ser utilizadas fontes de lipídeos predominantemente saturados (como a gordura de palma) ou insaturados (como os demais de origem vegetal, mas principalmente o de soja).
O efeito da suplementação de gordura protegida (na verdade, sais cálcicos de ácidos graxos) para bovinos leiteiros demonstra resultados variáveis sobre a composição do leite, dependendo da composição e da quantidade de gordura oferecida.
A suplementação de gorduras à base de ácidos graxos (AG) saturados é mais segura, pois estes são mais estáveis no rúmen, portanto contribuem para manutenção do teor de gordura no leite. Já as fontes ricas em AG polinsaturados são mais dependentes do pH ruminal para sua não-dissociação e consequente manutenção dos teores de gordura no leite.
No experimento conduzido por Carvalho et al. (2010), vacas suplementadas com sais cálcicos de ácidos graxos de óleo de palma produziram leite com maiores teores (3,26 vs. 3,11%) e quantidades (1,15 vs. 1,08 kg/dia) de gordura do que vacas suplementadas com gordura protegida de óleo de soja. Este efeito também foi observado por Relling & Reynolds (2007), os quais compararam a adição de gorduras inertes a base de AG saturados, AG monoinsaturados ou AG polinsaturados.
A explicação por esta moderada depressão da gordura do leite nos animais suplementados com sais cálcicos de AG de óleo de soja é que os AG insaturados encontrados nesta fonte de gordura supostamente inerte não são tão eficientemente protegidos contra a biohidrogenação parcial no rúmen, fato já demonstrado por Chouinard et al. (1998). Mudanças na biohidrogenação destes AG insaturados devem ter contribuído para a síntese do ácido linoléico conjugado trans-10 cis-12, potente inibidor da síntese de gordura do leite na glândula mamária (Bauman & Griinari, 2003).
Praticamente a totalidade dos experimentos consultados com suplementação de gorduras resultou em redução na porcentagem de proteína (Rabiee et al., 2012), principalmente da caseína. Esta redução pode ter impacto negativo nos atuais sistemas de pagamento do leite por qualidade. Práticas para amenizar esta redução nos teores proteicos do leite seria a suplementação com niacina, formulação de dietas com maiores níveis de PNDR (proteína não degradável no rúmen) e suplementação de aminoácidos protegidos ou análogos.
Somatotropina bovina
A condição nutricional das vacas tanto antes quanto durante a suplementação de somatotropina bovina (bST) determina o efeito deste hormônio sobre as concentrações de gordura e proteína no leite (NRC, 2001). Quando se faz uso de somatotropina bovina associada a uma alimentação e manejo adequados e em balanço energético positivo, as porcentagens de gordura, proteína, lactose, vitaminas e minerais não variam em animais suplementados (Raymond et al., 2009). Porém, quando tais requerimentos não são atendidos e o balanço energético negativo é prolongado, a porcentagem de gordura aumenta, a porcentagem de proteína diminui e a composição das gorduras do leite é alterada; com maior proporção de ácidos graxos de cadeia longa, oriundos da mobilização mais intensa das reservas corporais (Bauman et al., 1989).
Resumo das práticas nutricionais e de manejo que afetam a composição do leite
Na tabela 12 estão resumidas importantes práticas de nutrição e de manejo que podem afetar tanto a produção de leite, como os teores de gordura e proteína.

Conclusões
Nutrição, manejo e melhoramento genético são os principais meios de se alterar a composição do leite. Mudanças na composição do leite através da genética são obtidas a longo prazo quando comparadas às mudanças produzidas por práticas de nutrição e de manejo, que geralmente ocorrem de forma quase imediata. Por outro lado, as mudanças obtidas pela genética são permanentes, ao contrário das alterações proporcionadas pelas práticas de nutrição, que são transitórias.
Nutricionalmente, os teores de gordura e proteína no leite são antagônicos. Assim, práticas nutricionais e de manejo que aumentam a produção de leite normalmente também aumentam os teores de proteína do leite, mas por outro lado estas mesmas práticas diminuem os teores de gordura do leite.
Se você tem interesse em saber mais sobre a Nutrição dos Bovinos: Corte e Leite, te convido a conhecer a plataforma da AgricOnline. Ao fazer a sua assinatura, você tem acesso ilimitado a todos os cursos da plataforma. São cursos que vão desde produção vegetal, produção animal, mercado e carreira.
Ao término de cada curso, você tem direito ao certificado com a carga horária de cada curso, clique no link para conhecer.

Ou clique no link:
https://go.agriconline.com.br/pass/?sck=portal
Fonte
DE ALMEIDA, Gercílio Alves Júnior; STRADIOTTI Deolindo Júnior; DA SILVA, Elaine Cristina Gomes; ANDRADE; Magda Aparecida Nogueira; DE ALMEIDA, Maria Izabel Vieira; CÓSER, Antônio Carlos. Avanços Tecnológicos na Bovinocultura de Leite. 1ª ed. Alegre - ES: CAUFES, 2012.



