CONDIÇÃO CORPORAL E ATIVIDADE REPRODUTIVA DE ÉGUAS


A manutenção dos animais em boa condição corporal é de grande importância para se alcançar altos índices reprodutivos, tanto à monta natural quanto frente às modernas biotecnologias da reprodução equina. A presente revisão de literatura teve como objetivo abordar a influência da condição corporal sobre a atividade reprodutiva de éguas, enfocando a atividade ovariana, o perfil endócrino, sendo dada ênfase à leptina, pois seus níveis estão diretamente relacionados à condição corporal e envolvidos no estímulo do eixo hipotálamo-hipófise-gônadas, e os índices de eficiência reprodutiva. Considerando uma escala de escore da condição corporal (1 a 9), éguas com escore > 5 são consideradas aptas a apresentarem índices de eficiência reprodutiva satisfatórios. Em síntese, a condição corporal exerce efeito sobre: a duração do anestro estacional e do período interovulatório, a secreção de IGF-1 e leptina, o número de ciclos/concepção, as taxas de prenhez, perda embrionária precoce e aborto.

INTRODUÇÃO
Em um programa reprodutivo de equinos espera-se o nascimento de um potro saudável por égua, dentro do intervalo de um ano. Para isso, é necessário que se tenha éguas e garanhões de boa fertilidade, além de manejos reprodutivo, nutricional e sanitário adequados.
Para a avaliação da fertilidade são adotados índices de eficiência reprodutiva, que refletem a fertilidade tanto individual como do plantel. Vários estudos vêm sendo realizados visando o aumento da eficiência reprodutiva das éguas, alguns deles com o uso de fármacos (Thompson e Witherspoon, 1974; Alcântara et al., 2005; Meyers-Brown et al., 2013) e de biotecnologias avançadas da reprodução equina (Coutinho da Silva et al., 2004; Meyers-Brown et al., 2011; Nagao et al., 2012). Porém, de nada vale a utilização dessas ferramentas e inovações, sem que se tenha um plantel em boa condição corporal.
Resultados de pesquisas comprovam que éguas em boa condição corporal apresentam uma maior eficiência reprodutiva, quando comparadas àquelas em má condição corporal (Gentry et al., 2002ab; Gastal et al., 2004;), destacando sua importância na reprodução. A presente revisão de literatura teve por objetivo abordar a influência da condição corporal sobre a atividade reprodutiva de éguas.
DESENVOLVIMENTO
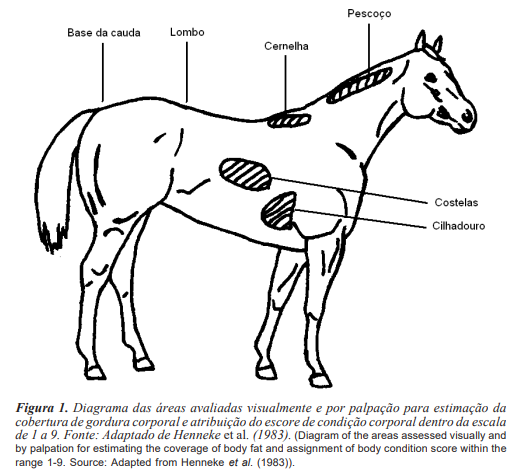
A condição corporal é determinada pela quantidade de gordura que um animal possui. Para estimá-la pesquisadores desenvolveram diferentes técnicas, buscando sempre maior confiabilidade e precisão (Henneke et al., 1983; Gentry et al., 2004; Carter et al., 2009). Henneke et al. (1983) definiram um sistema de escore da condição corporal (ECC) baseado na visualização e palpação de seis áreas corporais em que ocorre a deposição de gordura, sendo elas: bordo dorsal do pescoço, garrote, lombo, base da cauda, costado e a ligação da espádua com o costado (figura 1). Após a avaliação atribui-se um escore considerando uma escala de 1 a 9, sendo escore 1 o animal emaciado e escore 9 aquele extremamente obeso. Dentre as técnicas estabelecidas, esta é a mais aceita, sendo utilizada na experimentação em equinos avaliando, por exemplo, o efeito da condição corporal sobre o desenvolvimento folicular (Gastal et al., 2004) e as concentrações circulantes de leptina (Waller et al., 2006).
Gentry et al. (2004) estabeleceram uma nova forma de mensurar a gordura subcutânea utilizando a ultrassonografia. Tendo como base a técnica estabelecida por Henneke et al. (1983), os autores concluíram que a base da cauda é a melhor área para se determinar o acúmulo de gordura e, assim, definir o ECC. Esta é uma técnica de mensuração objetiva, no entanto apresenta um maior custo, pois necessita de um equipamento de ultrassom (Vecchi et al., 2010).
Manso Filho et al. (2009) avaliaram, pela ultrassonografia da região da garupa, a percentagem de gordura corporal de animais pertencentes a diferentes categorias e estágio fisiológico, criados em região tropical, e obtiveram valores iguais a 10,4; 14,0; 14,8; 15,5; 12,58 e 11,46 % para potras (<18 meses), éguas vazias, nos 2/3 iniciais de gestação, no 1/3 final de gestação, 1/2 inicial da lactação e 1/2 final da lactação, respectivamente. Os autores destacam que a determinação da proporção de gordura corporal tem importante função no entendimento das adaptações metabólicas durante as diferentes fases produtivas e reprodutivas, o que permite a avaliação e o estabelecimento de práticas de manejo nutricional adequadas às diferentes exigências.
A avaliação da condição corporal não depende do tamanho do animal, podendo ser utilizada inclusive em pôneis (Carter et al., 2009). A presença de pelos mais longos e espessos durante o período de inverno dificulta a visualização das áreas em que ocorre a deposição de gordura, devendo nestes casos se dar atenção especial para a palpação das mesmas (Henneke et al., 1983).
Dependendo da conformação corporal e estágio fisiológico dos animais, a avaliação deve ser adaptada, ou seja, as áreas a serem visualmente avaliadas não devem ser influenciadas por estes fatores. Em éguas que possuem garrote mais proeminente e maior deposição de gordura na região do lombo, deve-se dar ênfase para a avaliação do costado, ligação da espádua com o costado e base da cauda. No caso das éguas gestantes, à medida que elas se aproximam do parto, a pele e a musculatura da região dorso-lombar, costado e da base da cauda se distendem, diminuindo a cobertura de gordura nessas áreas, portanto, deve-se focar nos depósitos de gordura atrás das espáduas e no garrote. Éguas jovens não gestantes, aparentemente, possuem maior cobertura de gordura no costado, quando comparadas às éguas velhas com mesma condição corporal, portanto, esta região não deve ser levada em consideração durante a avaliação (Henneke et al., 1983).
O ECC tem correlação positiva com a percentagem de gordura corporal, mas não possui associação com o peso, altura e o perímetro torácico (Henneke et al., 1983). Porém, quando a altura é semelhante entre animais de um mesmo plantel, o peso tem alta correlação (r= 0,70; p<0,001) com a condição corporal (Carrol e Huntington, 1988).
Dentre as formas disponíveis para aferição do peso, dois métodos são frequentemente usados: a balança para uma avaliação precisa e a fita de peso que estima o peso do animal por seu perímetro torácico (Carrol e Huntington, 1988; Wagner e Tyler, 2011). Porém, as fitas de peso vendidas comercialmente têm sua fórmula validada pelos fabricantes, o que pode causar uma variação à aferição, entre as diferentes marcas (Wagner e Tyler, 2011).
Wagner e Tyler (2011) compararam a acurácia da estimativa do peso corporal utilizando a fita e a fórmula e constataram que quando a primeira foi utilizada, o peso real foi subestimado, em média, em 65,81 kg, enquanto com a fórmula houve uma diferença média de 17,25 kg, portanto sendo esta considerada mais confiável. Idealmente, seria obter uma fórmula para cada raça, pois o peso é estimado pela fórmula que se utiliza dos dados do comprimento do corpo e o perímetro torácico, sendo: Peso corporal (kg) = (perímetro torácico2 (cm) x comprimento do corpo (cm)) / 11 880 (Carrol e Huntington, 1988; Wagner e Tyler, 2011).
CONDIÇÃO CORPORAL E PERFIL ENDÓCRINO
Gastal et al. (2004) relatam que o ECC das éguas não influenciou a taxa de crescimento do folículo ovulatório e os níveis séricos de FSH e LH. Contudo, Gentry et al. (2002a) analisaram a resposta da hipófise quanto a secreção do LH e FSH frente ao tratamento com GnRH e obser-varam que os níveis de LH foram maiores nas éguas em boa condição corporal, enquanto o FSH não apresentou diferença significativa entre os grupos. No entanto, as avaliações foram feitas no período de inverno, quando as éguas em má condição corporal estavam em anestro estacional, enquanto 75 % das éguas do grupo com boa condição corporal continuaram ciclando, o que pode explicar a diferença observada entre os grupos estudados. Os autores também detectaram níveis menores de progesterona nas éguas em má condição corporal, durante a estação anovulatória, o que condiz com a ocorrência de anestro.
A seleção do folículo dominante e ocorrência da ovulação são influenciadas pelo fator de crescimento semelhante à insulina - 1 (insulin-like growth factor-1; IGF-1), pois este desempenha um importante papel intrafolicular durante esses processos ovarianos (Ginther et al., 2004). De fato, éguas com maior ECC apresentaram níveis superiores de IGF-1 (Cavinder et al., 2007; Peugnet et al., 2010). Peugnet et al. (2010) observaram que as concentrações de IGF-1, nas éguas em má condição corporal e tratadas com hormônio do crescimento (GH), apresentaram um aumento mais lento dos níveis de IGF-1 quando comparadas às éguas em boa condição corporal, porém alcançaram um mesmo platô, apresentando um período interovulatório semelhante entre os grupos. Nas éguas em alta condição corporal, o tratamento com o GH não teve efeito significativo sobre a duração do período interovulatório.
Cavinder et al. (2007), trabalhando com éguas no pós-parto, em condição corporal moderada (escores 5-6) e alta (escores 7-8), observaram diferença significativa nos níveis séricos de LH, 13 dias após a primeira ovulação, os quais foram mais elevados nas éguas com ECC moderado. Apesar disto, os níveis de progesterona plasmática e os períodos de ovulação, detectados por ultrassonografia, não diferiram entre os grupos e os autores concluíram que éguas com ECC entre 5 e 6 são tão eficientes, de acordo com a taxa de gestação ao segundo cio pósparto, quanto àquelas com alta condição corporal.
Da mesma forma, um período curto de restrição alimentar (48 horas) não provocou perda de condição corporal, nem foi acompanhado de mudanças na atividade do eixo reprodutivo, portanto, sem mudanças significativas nas concentrações de FSH, LH e prolactina (McManus e Fitzgerald, 2000; Gastal et al., 2010).
Recentemente, Anderson et al. (2013) observaram que o grupo de éguas em má condição corporal (escore 3,3 ± 0,3; escala de 0 a 9), durante o terço final da gestação, apresentaram resistência à insulina, o que não foi acompanhado pelo aumento da produção do hormônio. Estes resultados caracterizam um possível diabetes mellitus gestacional, que pode estar relacionada com a condição corporal da égua.
Assim, quando comparadas éguas em boa e má condição corporal, a diferença nos níveis séricos de LH e IGF-1 parece não influenciar a atividade ovariana quando as éguas com baixo ECC não se apresentarem em anestro.
LEPTINA, CONDIÇÃO CORPORAL E ATIVIDADE REPRODUTIVA
A leptina é uma proteína de 16-kDa secretada pelos adipócitos e tem importante função na regulação do balanço energético e metabolismo. Receptores para leptina são encontrados em diversos tecidos como fígado, rins, coração, músculo esquelético e pâncreas, bem como no hipotálamo e adenohipófise, tendo funções fisiológicas importantes na modulação da sensibilidade à insulina e na regulação das funções imunológica e reprodutiva (Radin et al., 2009). A leptina vem sendo cada vez mais estudada, pois seus níveis estão diretamente relacionados à condição corporal e envolvidos na estimulação da função hipotalâmica-hipofisária-gonadal (Buff et al., 2002; Ferreira-Dias et al., 2005; Huff et al., 2008). Em equinos a leptina tem um padrão circadiano de secreção, com picos ocorrendo durante o período noturno, e, diferindo do observado em humanos, éguas apresentam níveis significativamente mais elevados de leptina circulante que os garanhões e machos castrados (Radin et al., 2009).
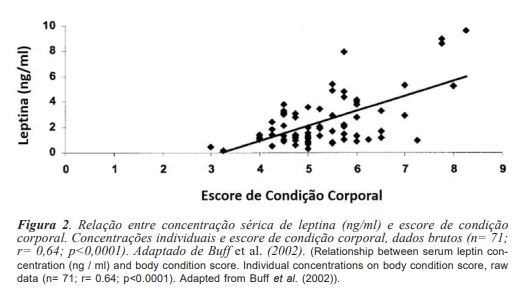
O período anovulatório é precedido por uma queda nos níveis de leptina (Fitzgerald e McManus, 2000; Ferreira-Dias et al., 2005), mesmo quando a condição corporal é mantida (Waller et al., 2006). Os níveis de leptina plasmática têm correlação significativa com o ECC (r= 0,64; figura 2 - Buff et al., 2002). Éguas com bom ECC apresentam período anovulatório menor e níveis mais altos de leptina, retornando a atividade ovariana antes do equinócio de primavera, enquanto éguas com menores níveis de leptina reiniciam a atividade cíclica mais tarde, já na primavera, e este reinício da atividade ovariana é acompanhado por um aumento da concentração plasmática de leptina (Fitzgerald e McManus, 2000; Ferreira Dias et al., 2005).
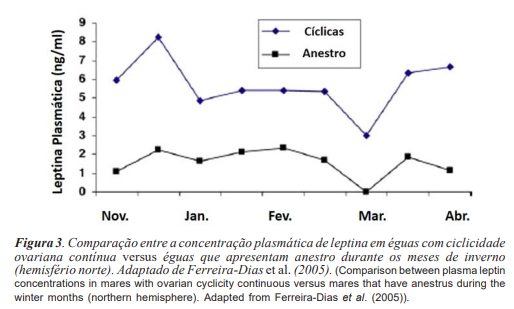
As fêmeas que mantêm níveis altos de leptina durante o inverno podem chegar a apresentar atividade ovariana cíclica durante o ano todo, enquanto as éguas que apresentam baixos níveis de leptina (< 3 ng / ml) durante o inverno tendem a ser acíclicas neste período (figura 3 - Ferreira-Dias et al., 2005; Èebulj-Kadunc et al., 2009). No entanto, Waller et al. (2006) não observaram esta influência sobre a manutenção da ciclicidade, atribuindo este fato somente ao bom ECC das éguas. Possivelmente, este contraste entre resultados se deveu ao fato de no experimento de Waller et al. (2006), as éguas consideradas com baixos níveis de leptina apresentaram concentrações variando de 3 a 5 ng / ml. Isto leva à hipótese de que apenas níveis abaixo de 3 ng / ml de leptina afetam negativamente a atividade ovariana de éguas, com aumento do período anovulatório.
A variação sérica das concentrações de leptina foi observada no decorrer do ano, sendo que durante a primavera e verão, período que as éguas ganharam peso, houve um aumento dos níveis de leptina enquanto que no outono e inverno foi observado o inverso, uma diminuição do peso corporal com uma queda da concentração circulante de leptina (Fitzgerald e McManus, 2000). No entanto, Walter et al. (2006) relataram que parece existir uma variação sazonal da leptina sem uma mudança correspondente da condição corporal, pois a redução plasmática de leptina foi observada durante o inverno, mesmo em éguas que mantiveram alta condição corporal.
Da mesma forma, há uma grande variação na concentração plasmática de leptina entre éguas com alto ECC, indicando que outros fatores podem estar envolvidos na sua síntese e secreção (Gentry et al., 2002a; Huff et al., 2008). Portanto, além do perfil sazonal a concentração de leptina pode apresentar uma variação individual. Porém, esta parece não afetar o perfil endócrino da atividade ovariana, pois éguas que apresentaram altos ou baixos níveis médios de leptina no plasma (>10 e <5 ng/ml, respectivamente), não diferiram entre si quanto as concentrações de FSH, LH, progesterona (Waller et al. 2006) e estradiol (Romagnoli et al., 2007). Desta forma, Waller et al. (2006) acreditam que a atividade ovariana esteja mais relacionada à condição corporal do que propriamente com a disponibilidade de leptina. Huff et al. (2008) observaram que a hiperleptinemia (>10 ng/ml) tende a se apresentar em éguas com alto ECC, porém, o mesmo pode ser observado quando em baixo ECC, o que levou os autores a supor que um fator genético possa estar envolvido.
A idade também exerce efeito sobre os níveis circulantes de leptina, pois animais jovens tendem a ter menores valores de leptina sérica quando comparados aos adultos, podendo este fato ser atribuído à menor deposição de tecido adiposo, devido ao desenvolvimento corporal. Potras de 2 anos de idade apresentaram menores concentrações de leptina, que àquelas de 3 e 4 anos (Èebulj-Kadunc et al., 2009), enquanto os animais com idade entre 5 e 12 anos apresentaram níveis de leptina mais altos, porém, as maiores concentrações foram detectadas em animais acima de 12 anos, talvez pela menor atividade dos mesmos (Buff et al., 2002).
Há relato de que a utilização de substâncias que reduzem o percentual de gordura corporal, como o clembuterol, provoca uma concomitante diminuição dos níveis circulantes de leptina (McManus e Fitzgerald, 2003).
A restrição alimentar de curto prazo, sem alteração da condição corporal, também pode influenciar a concentração de leptina plasmática (McManus e Fitzgerald, 2000; Gastal et al., 2010). McManus e Fitzgerald (2000) suspenderam o alimento de éguas em boa condição corporal por 24 horas e observaram que a diminuição da leptina tornou-se significativa após 11 horas de restrição, e esses níveis mantiveram-se baixos até 6 horas depois do retorno à alimentação. Os referidos autores também dosaram as concentrações de glicose e ácidos graxos livres e verificaram que a glicose se mostrou estável durante a restrição alimentar, enquanto os ácidos graxos tiveram um aumento significativo de seus níveis. Com isso os autores levantaram a hipótese de que níveis altos ou baixos de leptina não podem, por si só, serem determinantes da atividade reprodutiva, pois se houver substrato metabólico suficiente na circulação ou para mobilização, pode não ocorrer uma supressão do eixo reprodutivo. Neste mesmo sentido, as éguas que passaram por longos períodos de restrição alimentar, com redução da condição corporal, tendo baixos níveis de leptina e possuindo reduzido estoque de substrato metabólico de reserva, podem, supostamente, apresentar sua atividade reprodutiva comprometida. Gastal et al. (2010) constataram, após submeterem as éguas a um período de 48 horas de restrição alimentar, que a diminuição da concentração de leptina plasmática pode chegar a aproximadamente 55 %.
Éguas Lusitanas em regime extensivo de criação, com ECC > 2,25 (escala 1 – 5) apresentaram aumento das concentrações plasmáticas de leptina no terço final de gestação e queda desses níveis durante o primeiro mês de lactação, porém esta variação não teve efeito sobre o intervalo do parto à concepção, e os resultados de fertilidade ao 1º e 2º ciclos estrais pós-parto (Fradinho et al., 2014). Éguas com ECC moderado (5-6) e alto (7-8), a primeira e segunda ovulação pós-parto, tiveram valores médios similares de leptina durante o ciclo estral, bem como de eficiência reprodutiva (Cavinder et al., 2007). A concentração de leptina sérica diminuiu entre 2 semanas antes, até 10 semanas após o parto, quando foi observado um concomitante aumento da concentração de leptina no leite, sendo seu maior nível obtido no momento do parto. A concentração de leptina se manteve baixa até 21 semanas pós-parto (Romagnoli et al., 2007) e o metabolismo da leptina, no potro recém-nascido, parece não depender da condição corporal e do metabolismo lipídico da mãe (Kêdzierski et al., 2011).
Os efeitos da leptina sobre a atividade reprodutiva ainda não estão totalmente esclarecidos, mesmo assim é possível observar que existe variação circadiana, sazonal, individual e durante o ciclo estral dos níveis plasmáticos desse hormônio. Porém, níveis de leptina <3 ng/ml associados a uma diminuição da reserva energética parece sinalizar para uma redução da atividade reprodutiva. Esses efeitos ainda precisam ser mais explorados, servindo como base para futuros estudos.
CONDIÇÃO CORPORAL E ATIVIDADE OVARIANA
A atividade ovariana cíclica das éguas utilizadas na reprodução é de grande importância para que se obtenham bons índices reprodutivos. Diversos autores constataram que a função ovariana pode ser influenciada pela condição corporal, sendo menor em éguas que apresentam um baixo ECC (Henneke et al., 1984; Gentry et al., 2002ab; Gastal et al., 2004; Peugnet et al., 2010).
Éguas em má condição corporal não apresentam atividade folicular significativa e permanecem em anestro durante 6 a 7 meses do ano. Algumas destas éguas podem inclusive demonstrar sinais comportamentais de estro, tais como a abertura dos membros posteriores, micção e eversão do clitóris, quando expostas ao garanhão. No entanto, éguas em boa condição corporal mantêm alguma atividade ovariana e a maioria delas apresentam ciclos estrais durante os meses de inverno (Gentry et al., 2002ab; Ferreira-Dias et al., 2005; Vecchi et al., 2010; Salazar-Ortiz et al., 2011).
Após o término do anestro estacional segue-se o período de transição para a atividade ovariana cíclica, o qual também pode ser influenciado pelo ECC das éguas. Gastal et al. (2004) observaram maior incidência de ondas anovulatórias nas éguas em má condição corporal (3/7), que naquelas com boa condição corporal (0/6), assim, o período de transição foi maior para as éguas em má condição corporal. Não foi observada diferença significativa entre os grupos quanto à duração do primeiro período interovulatório.
Por outro lado, resultados de pesquisa indicam que no decorrer da estação de monta houve influência do ECC sobre o período interovulatório. Henneke et al. (1984) relatam diferença significativa, quanto a duração média do período interovulatório, entre éguas que mantiveram ECC < 5 (31 dias), e aquelas com ECC > 5, ou que iniciaram a estação reprodutiva com baixo ECC, mas, que estavam ganhando condição corporal durante a estação de monta (22,7 dias). Peugnet et al. (2010) obtiveram resultados semelhantes, com as éguas em má condição corporal apresentando um intervalo médio entre ovulações de 28,4 dias, enquanto que para aquelas com boa condição corporal foi de 23 dias.
Alguns autores relataram o efeito do ECC sobre o desenvolvimento do folículo ovulatório, demonstrando que éguas em má condição corporal apresentam um diâmetro do folículo pré-ovulatório significativamente menor quando comparado com o de éguas em boa condição corporal (Godoi et al., 2002; Gastal et al., 2004; Rodrigues et al., 2011). Nesse contexto, Gastal et al. (2010) avaliaram éguas após um curto período de restrição alimentar (48 horas) e observaram, pela ultrassonografia doppler, que as mesmas possuíam maior vascularização do folículo pré-ovulatório, sugerindo que esta talvez seja uma resposta compensatória do organismo para proteger as células foliculares da hipóxia e apoptose. Avaliando éguas doadoras de embrião, Rodrigues et al. (2011) não observaram influência da condição corporal sobre a taxa de recuperação de embriões. Entretanto, o maior diâmetro do folículo ovulatório foi associado a uma maior taxa de recuperação embrionária (p<0,05), o que segundo os autores demonstra uma influência indireta da condição corporal sobre este índice.
Considerando o intervalo entre o parto e a primeira ovulação, a condição corporal não teve efeito sobre esta característica, mas tendeu a ser maior para as éguas com ECC < 5 (16,8 vs. 12,7 dias - Henneke et al., 1984). De fato, Godoi et al. (2002) obtiveram uma correlação negativa entre estas variáveis (r = -0,50; p<0,02).
De acordo com os resultados apresentados, a má condição corporal pode levar a queda dos índices reprodutivos, em decorrência da diminuição da atividade ovariana e do tamanho do folículo ovulatório, e aumento do período de anestro estacional, da duração do período de transição para a retomada da atividade cíclica e do intervalo interovulatório. A associação desses fatores pode levar a prejuízos econômicos diretos em uma propriedade, bem como resultar no nascimento dos potros em um período menos favorável para a comercialização.
CONDIÇÃO CORPORAL E EFICIÊNCIA REPRODUTIVA
O número de ciclos/concepção é um índice que permite realizar o diagnóstico precoce de problemas individuais ou daqueles envolvendo o manejo reprodutivo geral do plantel. Henneke et al. (1984) observaram que éguas que iniciaram a estação de monta com ECC < 5 necessitaram em média de 3,1 ciclos/concepção, enquanto aquelas com ECC > 5, requereram 1,43 ciclos/concepção. A taxa de gestação de éguas em lactação não diferiu ao 1º ciclo, já ao 2º e 3º ciclos a diferença foi significativa (ECC < 5 = 50%; ECC > 5 = 100 %).
Em um segundo experimento, Henneke et al. (1984) avaliando éguas virgens, vazias e paridas, constataram que a taxa de gestação ao final da estação de monta diferiu significativamente entre éguas com ECC inferior e superior a 5 (66 % e 95,43 %, respectivamente), resultado também relatado por Zúccari et al. (2013), em que éguas com ECC < 3,5 e > 5 (escala de 1 a 9), tiveram taxa de gestação de 7,1 % e 63,6 % (p<0,05), respectivamente. Além disso, a taxa de aborto, avaliada aos 90 dias de gestação, também apresentou diferença significativa entre os grupos, sendo esta maior quando ECC<5 (75 % vs. 0 %, respectivamente; Henneke et al.,1984); contudo, vale ressaltar o pequeno tamanho amostral, sendo acompanhadas apenas quatro éguas, e três delas apresentaram aborto. Newcombe e Wilson (2005) observaram que éguas que estão perdendo peso durante a concepção e gestação, têm maiores chances de apresentar perda embrionária precoce. Dentro desse grupo de animais (n= 20), 75 % (15 / 20) apresentaram perda embrionária até os 32 dias de gestação e 66,67 % (8 /12) entre os dias 29 a 45. No mesmo sentido, Miyakoshi et al. (2012) demostraram, com avaliação realizada aos 35 dias de gestação, diferença significativa na taxa de perda embrionária precoce, sendo esta maior quando as éguas estavam com ECC < 5 (11,8 % vs. 3,8 %), e, da mesma forma, também apresentaram maior taxa de aborto (23,4 % vs. 7,6 %).
A obesidade pode interferir negativamente na eficiência reprodutiva de algumas espécies. No entanto, o mesmo não foi observado em éguas por Kubiak et al. (1989). Os autores verificaram que grupos de éguas paridas pluríparas, mantidas com ECC entre 5,5 e 7 (grupo controle) ou igual a 9 (obesas), não diferiram quanto ao número de fêmeas em cio ou na intensidade dos sinais e duração do cio. Da mesma forma, não houve diferença significativa entre os grupos para o intervalo entre o parto e 1º e 2º cios, número de ciclos/concepção e taxa de gestação ao 1º ciclo. Todas as éguas tornaram-se gestantes e não houve perda da prenhez.
Devido à longa duração da gestação dos equinos, o período de serviço deve ser o menor possível, visando um ótimo intervalo entre partos. Portanto, o manejo no período pós-parto é um dos fatores de grande importância em uma criação. Nagy et al. (1998) acompanharam cinco plantéis, durante duas estações de monta consecutivas, e constataram uma tendência, das éguas com perda da condição corporal pós-parto, apresentarem um maior intervalo entre a primeira e segunda ovulações pós-parto (p= 0,069) e uma duração mais longa da fase folicular no segundo ciclo estral (p= 0,089).
O período de gestação e o peso do potro ao nascimento não parecem ser influenciados pela condição corporal, sendo em média de 344 dias e 50 Kg, respectivamente para éguas Quarto de Milha. Estes dados dão suporte à teoria de que o feto tem prioridade sobre os nutrientes presentes na circulação sanguínea materna e, no caso de dieta limitada, estes são mobilizados a partir de reservas metabólicas, para o adequado crescimento e desenvolvimento do feto (Henneke et al., 1984). A taxa de potros vivos ao nascimento também não depende da condição corporal, não havendo diferença significativa entre grupos com alta ou baixa condição corporal (Cameron et al., 1999). Henneke et al. (1984) ao analisarem peso, altura e perímetro torácico de potros aos 90 dias de idade, não constataram diferença significativa quanto ao desenvolvimento corporal, portanto, o mesmo não sofreu influência da condição corporal das éguas. Aparentemente, o grupo de éguas que manteve ECC < 5 utilizou energia de reserva para a produção de leite. Por outo lado, Fradinho et al. (2014) relatam que a perda de ECC das éguas, no primeiro trimestre da lactação, teve efeito negativo significativo sobre a taxa de crescimento de seus potros.
Kubiak et al. (1988) não observaram efeito da obesidade (ECC = 8,5 – 9) sobre a parição e peso do potro ao nascer, no entanto, em estudo posterior foi observada uma tendência de éguas obesas apresentarem uma menor produção de leite, com potros significativamente mais leves e com menor perímetro torácico aos 63 dias pósparto. Diversas hipóteses foram levantadas para tentar explicar estes resultados, como maior deposição de gordura nas glândulas mamárias, nascimento de potros menos vigorosos ou com menor potencial de crescimento, com consequente redução da demanda de leite, não podendo também ser descartado o fator genético (Kubiak et al., 1991).
Observações em humanos e animais experimentais têm demonstrado que o baixo peso ao nascer, decorrente de uma má nutrição do feto durante a gestação, está ligado à disfunção dos sistemas cardiovascular, metabólico e endócrino na vida adulta. Uma das causas do fornecimento inadequado de nutrientes ao feto é o estado nutricional materno. Essas mudanças permanentes na estrutura e função dos tecidos, causadas por injúrias durante a vida fetal, se denominam programação intrauterina (Mcmillen e Robinson, 2005; Knight et al., 2009; Van Abeelen et al., 2012; Fowden et al., 2013). No entanto, pouco se sabe sobre a programação intrauterina em animais domésticos de vida longa. Fowden et al. (2013) levantaram a hipótese que este efeito pode estar presente também nos equinos, porém uma área que ainda necessita de estudos.
Após o primeiro terço da gestação, a diminuição do ECC parece não ter grande influência no desenvolvimento do concepto, porém, este é um fator que não deve ser negligenciado, já que logo após o parto a égua irá destinar boa parte de suas reservas energéticas à lactação, podendo haver uma perda ainda maior da condição corporal. Além disso, a má condição no pós-parto acarreta perdas econômicas, tendo em vista que levará a um aumento do número de ciclos por concepção, da perda embrionária precoce e do intervalo entre partos.
CONCLUSÕES
A eficiência reprodutiva está intimamente relacionada com a condição corporal, a qual exerce efeito sobre a duração do anestro estacional, do período de transição para a atividade cíclica, do período interovulatório, a secreção de IGF-1 e leptina, o número de ciclos/concepção e as taxas de gestação, de perda embrionária precoce e de aborto. Desta forma, constata-se que a manutenção das éguas em condição corporal adequada, ECC > 5 (escala de 1 a 9), é um pré-requisito a ser atendido para que os índices de eficiência reprodutiva sejam satisfatórios. Nos sistemas de produção de equinos devem ser desenvolvidas estratégias alimentares que visem impedir a queda do ECC das éguas, para que as perdas econômicas sejam reduzidas. De forma geral, a utilização do escore de condição corporal se mostra uma ferramenta útil para a avaliação e predição da eficiência reprodutiva de um rebanho equino. Os efeitos da leptina sobre a atividade reprodutiva dos equinos ainda precisam ser mais explorados quanto às variações individuais, circadianas, sazonais e durante o ciclo estral, além de seu papel à puberdade. Pesquisas são necessárias para se avaliar qual a concentração mínima de leptina circulante para o estabelecimento da atividade ovariana, buscando-se validá-la como um possível indicador do potencial de ciclicidade das éguas.
Fonte
BENDER, C; SAMPAIO, B; NOGUEIRA, B G; et al. Condição corporal e atividade reprodutiva de eguas. Archivos de Zootecnia, v. 63, n. 241, p. 55–67, 2021. Disponível em: <https://www.uco.es/servicios/ucopress/az/index.php/az/article/view/590/564>. Acesso em: 24 out. 2022.



