Cinética de absorção e frações nitrogenadas em arroz expressando o fator de transcrição OsDof261


Fatores de transcrição Dof podem atuar como ativadores ou repressores transcricionais envolvidos em diversos processos biológicos exclusivos de vegetais, no entanto muitas de suas funções fisiológicas ainda não foram elucidadas. Neste trabalho, plantas de arroz (Oryza sativa L.) expressando o fator de transcrição OsDof26 mostraram alterações na cinética de absorção e metabolismo de nitrogênio (N) quando submetidas aos tratamentos de 0,2 e 2,0 mM de amônio (NH4+) ou nitrato (NO3-) em solução nutritiva. O delineamento experimental foi inteiramente casualizado, com quatro repetições, em esquema fatorial 2 × 2 × 2, duas plantas (plantas de arroz não transformadas e uma linhagem de arroz expressando oOsDof26, a L24, duas fontes de N (NO3- e NH4+) e duas doses (0,2 e 2,0 mM). Aos 17 dias após a germinação (DAG) foram aplicados os tratamentos e procedeu-se a coleta da solução nutritiva em intervalos de 30 minutos até as 48 horas, afim de estimar os parâmetros cinéticos de absorção de N (Vmax, Km e Cmín). Aos 19 DAG, obteve-se a massa fresca total, sendo parte desse material armazenado para determinação das frações nitrogenadas e açúcares solúveis. A L24 apresentou menor crescimento da parte aérea e raiz em todos os tratamentos analisados quando comparada com a planta não transformada (WT -wild type). e menores teores de NO3-, NH4+ e N-amino, afetando assim o metabolismo de N nas plantas transformadas.
INTRODUÇÃO
O nitrogênio (N) é o nutriente que mais limita a produtividade dos cultivos e o teor de proteína nos grãos de cereais (DECHORGNAT et al., 2011). Diante disso, a eficiência de uso de N (EUN) é considerada crítica para o crescimento, ganho de biomassa, rendimento dos cultivos, bem como a ótima utilização dos fertilizantes aplicados (SHRAWAT; GOOD, 2008).
A EUN pode ser dividida em dois processos: eficiência de absorção, ou seja, habilidade da planta de retirar o N do solo, normalmente presente sob as formas de NO3- e NH4+, e eficiência de utilização, que é a habilidade da planta de transferir o N para o grão, predominantemente como proteína (LEA; AZEVEDO, 2006).
Duas estratégias podem ser usadas visando o aumento da eficiência de uso de nutrientes: uma envolve o melhoramento convencional e a seleção assistida por marcadores moleculares, numa tentativa de identificar os genes envolvidos, enquanto a outra utiliza construções genéticas para transformar as plantas e provê-las de ferramentas específicas quanto à eficiência de uso de nutrientes (MCALLISTER; BEATTY; GOOD, 2012).
A eficiência de uso de N é uma característica controlada por múltiplos genes, deste modo, são promissores os estudos de transformação gênica com proteínas regulatórias, também chamadas de fatores de transcrição. Fatores de transcrição Dof (DNA one finger) são exclusivos de plantas e representam uma classe particular de domínio de ligação ao DNA tipo dedo de zinco. Membros desta família têm sido encontrados assumindo diversas funções na regulação de genes e processos específicos das plantas (NOGUERO et al., 2013).
Análises bioquímicas e genéticas revelaram que proteínas de domínio Dof funcionam como ativadores ou repressores transcricionais envolvidos em diversos processos biológicos em vegetais. Embora muitas funções das proteínas Dof ainda não tenham sido elucidadas, é evidente que esses fatores desempenham funções críticas na regulação transcricional relacionada ao desenvolvimento das plantas (YANAGISAWAet al., 2004).
Plantas de Arabidopsis superexpressando o gene Dof1 de milho mostraram aumento na produção de esqueletos de carbono (C) e na assimilação do N absorvido, bem como melhor desempenho sob condições de baixa disponibilidade deste nutriente (YANAGISAWA et al., 2004). Por outro lado, estes mesmos autores relatam efeito inibitório da proteína Dof2 de milho sobre a Dof1 na expressão da enzima fosfoenolpiruvato carboxilase (PEPC), indicando uma possível ação conjunta destes dois fatores de transcrição no controle da expressão do gene da PEPC (YANAGISAWA; SHEEN, 1998).
Uma análise filogenética entre todos os membros desta família de fatores de transcrição em arroz mostra uma alta similaridade entre o OsDof25 e o OsDof26 (SANTOS; SOUZA; FERNANDES, 2012), indicando uma possível interação entre estes dois fatores de transcrição no controle da expressão gênica, a exemplo do que ocorre entre o Dof1 e Dof2 de milho.
Diante disso, este trabalho teve como objetivo avaliar se plantas de arroz transformadas expressando o fator de transcrição OsDof26 apresentam alterações na sua cinética de absorção e metabolismo de N quando submetidas a condições de baixa e alta disponibilidade de NH4+ ou NO3- na solução.
MATERIAL E MÉTODOS
A construção gênica para expressão do OsDof26 em plantas de arroz foi obtida por meio da tecnologia Gateway de clonagem (Invitrogen). O OsDof26 foi amplificado do genoma nuclear usando os seguintes iniciadores: 5’AAAAAGCAGGCTGCATGCAGGAGGCGGGGCGA-3’ e 5’- GGGGACCACTTTGTACAAGAAAGCTGGGT-3’.
O gene OsDof26 foi clonado no vetor pDONR221 (Invitrogen) usando o kit Gateway BP Clonase II Enzyme Mix seguindo as recomendações do fabricante. O vetor resultante pENTR-OsDof26 foi usado em uma recombinação múltipla com os vetores pENTR-UBIL, pENTR-3xHA e o vetor de expressão pH7m34GW, este último contendo o gene de resistência a higromicina usado na seleção das plantas transformadas. A reação de recombinação múltipla foi realizada usando o kit LR Conase II Plus Enzyme Mix (Invitrogen) seguindo as recomendações do fabricante.
O cassete de expressão obtido contém o gene OsDof26 sobre o controle do promotor forte da ubiquitina 1 de milho (UBIL:OsDof26:3xHA). A transformação de plantas de arroz foi realizada baseada no protocolo de Toki (2006). As linhagens foram cultivadas em casa de vegetação até a terceira geração para obtenção de plantas homozigotas.
O experimento foi conduzido em câmara de crescimento com radiação de 500 µmol fótons m2 s-1 (fluxo de fótons fotossintéticos), fotoperíodo de 14h/10h (luz/escuro), umidade relativa do ar de 80% e temperatura média de 28 ºC/24 ºC (diurna/ noturna). Foram utilizadas sementes da variedade de arroz Nipponbare (Oryza sativa L. ssp. Japonica) superexpressando o fator de transcrição OsDof26 (linhagem 24 - L24) e sementes de plantas não transformadas (WT) da mesma variedade.
As sementes foram primeiramente tratadas em solução de hipoclorito de sódio (2%) durante 10 minutos, depois lavadas repetidas vezes em água destilada e germinadas sobre gaze em vasos de 1,7 L contendo água destilada. Cinco dias após a germinação (DAG), 4 plântulas uniformes foram transferidas para potes de 700 mL contendo solução de Hoagland e Arnon (1950) modificada a ¼ força iônica (FI) com 2,0 mM de N (1,5 mM na forma de N-NO3- e 0,5 mM N-NH4+). Trocas de solução foram realizadas a cada três dias e o pH foi mantido em 5,7.
Aos 14 DAG as plântulas receberam solução a ½ FI sem N durante 72 horas, procedimento realizado para aumentar a capacidade de absorção do nutriente durante a cinética de absorção. Aos 17 DAG foram aplicados os tratamentos de 0,2 e 2,0 mM de N-NO3ou N-NH4+, sendo o NO3- fornecido na forma de KNO3 e o amônio de (NH4)2SO4. Plantas transformadas e não transformadas foram submetidas às mesmas soluções nutritivas. Foram realizadas coletas de 1,0 mL de solução nutritiva, sem reposição, em intervalos de 30 minutos para os tratamentos de 0,2 mM de N até às 24 horas e coletas em intervalos de 1 hora para os tratamentos de 2,0 mM de N até às 48 horas. Aos 19 DAG, as plantas foram separadas em parte aérea e raízes para determinação da massa fresca total.
Os parâmetros cinéticos de absorção (Influxo, Vmax e Km) de N-NO3- e N-NH4+ foram determinados a partir da diminuição da concentração destes íons na solução nutritiva em função do tempo, conforme descrito por Claassen e Barber (1974). O ajuste das curvas de depleção e os cálculos dos parâmetros cinéticos foram realizados empregando-se o software Cineticawin 1.0 (UFV). O poder de absorção (Į) foi calculado pela razão Į = Vmáx/Km e o Cmín foi estimado a partir das últimas coletas de solução nutritiva, quando os teores de N não sofreram variações.
Amostras de 0,5 g do material fresco coletado da parte aérea ou raízes foram homogeneizadas em etanol 80%, e após a extração alcoólica foi feita a partição com clorofórmio. A fração solúvel obtida foi utilizada para a determinação dos teores de N-amino livre (YEMM; COCKING, 1955), N-NO3- (CATALDO et al., 1975), N-NH4+ (FELKER, 1977) e açúcares solúveis (YEMM; WILLIS, 1954).
O delineamento experimental foi inteiramente casualizado em um fatorial (2 linhagens × 2 fontes de N × 2 doses) com quatro repetições. Os dados foram submetidos à análise de variância com aplicação do teste F e as médias comparadas entre si pelo teste de Tukey p 0,05 por meio do programa Sisvar para Windows versão 4.0 (FERREIRA, 2000) ou pelo erro padrão.
RESULTADOS E DISCUSSÃO
As plantas transformadas para superexpressão do fator de transcrição OsDof26 (L24) mostraram maior Km em todos os tratamentos analisados, com destaque para a dose de 0,2 mM de N-NO3- ou N-NH4+ (Tabela 1).
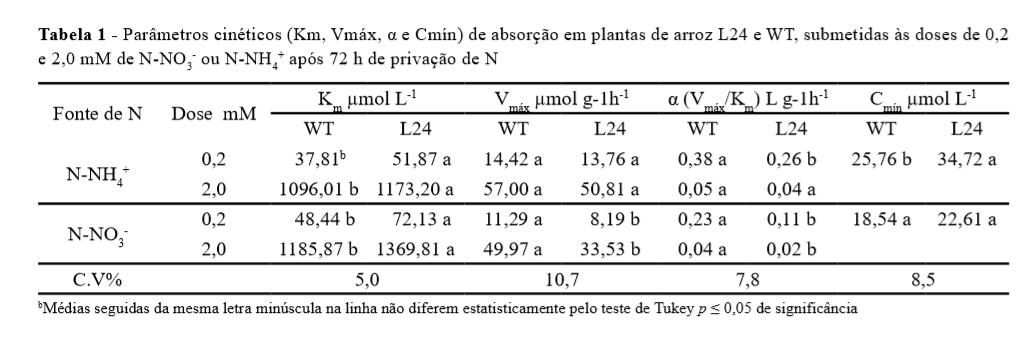
Por meio do valor de Km é possível verificar a afinidade dos carreadores pelo nutriente a ser transportado, sendo este o parâmetro mais relevante na descrição dos sistemas de transporte (ZHANG; RENGEL; MENEY, 2009). O maior Km observado na L24 representa uma perda de afinidade dos seus carreadores para o transporte de NO3- ou NH4+ presente na solução.
A Vmáx variou pouco entre L24 e WT (Tabela 1). Apenas nos tratamentos com N-NO3-, observou-se redução da Vmáx em L24 que pode ser reflexo de uma menor quantidade de proteínas de transporte para este ânion. Esta é uma característica desfavorável, uma vez que plantas que apresentam menor Vmáx mostram redução na capacidade de transporte de nutrientes (SANTOS et al., 2011).
Plantas de arroz transformadas para a superexpressão do fator de transcrição OsDof25, mostraram parâmetros cinéticos mais favoráveis (Vmáx e Km) em detrimento da WT quando submetidas a dose de 0,2 mM de NO3- ou NH4+ em solução nutritiva (SILVA, 2012).
Resultado contrário foi obtido em nosso trabalho, em que o aumento de expressão do OsDof26 gerou plantas com parâmetros cinéticos desfavoráveis em relação a WT tanto sob baixo, quanto alto suprimento de N (Tabela 1), indicando um possível efeito inibitório desse gene sobre o OsDof25.
A combinação de alto valor para Vmáx e baixos valores para Km e Cmín, juntamente com maior crescimento radicular, são características desejáveis numa espécie, pois representam maior eficiência na absorção do nutriente (LI et al., 2007). No entanto, plantas da L24 conduzidas com 0,2 ou 2,0 mM de N-NH4+ ou N-NO3mostraram maior Km e uma tendência de menor Vmáx. A análise conjunta dos parâmetros cinéticos por meio do poder de absorção (Į) corrobora a menor capacidade de absorção de N por L24 (Tabela 1).
O Cmín representa a concentração do nutriente em solução onde o influxo líquido é igual a zero e seu valor pode variar consideravelmente entre espécies de plantas e em diferentes condições ambientais (HU et al., 2010). Foram encontrados os valores de 25,76 e 34,72 µmol L-1 para a WT e L24, respectivamente. Este resultado, mais uma vez, indica que L24 mostrou menor capacidade de absorver o N da solução nutritiva.
Uma vez obtida a concentração do nutriente em solução a cada tempo de coleta, foram construídas curvas de influxo para os tratamentos de 0,2 e 2,0 mM de NNH4+ e N-NO3-. Foi observado maior influxo de N por WT (Figura 1).
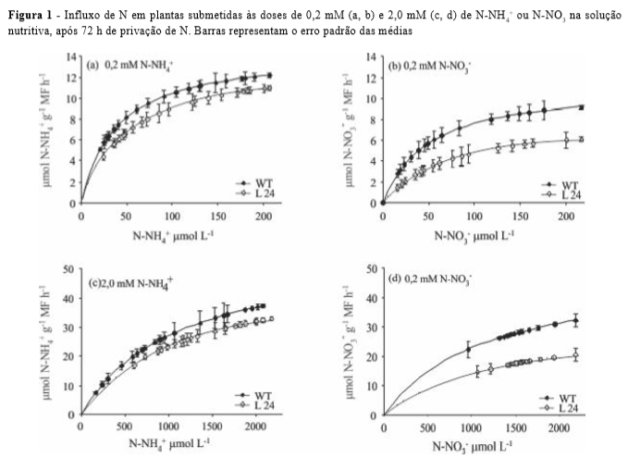
Estes dados corroboram os parâmetros cinéticos encontrados, em que L24 mostrou menor afinidade a NNH4+ ou N-NO3-, independente da dose de N.
Em todos os tratamentos, a L24 mostrou menor crescimento da parte aérea (Tabela 2).
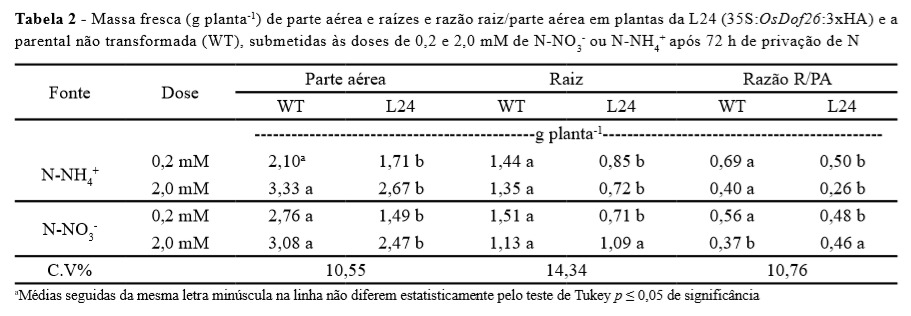
As raízes mostraram comportamento similar, no entanto, não foi observado diferença estatística no tratamento de 2,0 mM de N-NO3(Tabela 2). O aumento de expressão do OsDof26 gerou plantas com menor taxa de crescimento e com alterações na capacidade de absorção de N (Tabela 1, Figura 1).
A proteína Dof1 de milho em Arabidopsis pode atuar em funções críticas na regulação transcricional relacionada ao desenvolvimento das plantas, aumentando a expressão de genes relacionados à síntese de esqueletos de C, especialmente a piruvato quinase (PK) e PEPC (YANAGISAWAet al., 2004). Dados recentes mostraram que plantas de arroz superexpressando oOsDof25, ortólogo de Dof1 de milho, podem apresentar maior biomassa sob condições de estresse nutricional (deficiência ou baixo suprimento de N) (SILVA, 2012).
A superexpressão de OsDof26, provável ortólogo de Dof2 de milho, pode ter regulado negativamente a expressão de genes envolvidos na síntese de esqueletos de C que estão diretamente relacionados à assimilação de N. O menor crescimento de L24 sugere que a ocorrência de alterações no metabolismo de C e N podem ter prejudicado o desenvolvimento das plantas transformadas.
A razão raiz/parte aérea (R/PA) sofreu alterações entre as plantas estudadas (Tabela 2), onde foi observada uma tendência de menor valor da razão para a L24 o que estaria associado a um menor crescimento do sistema radicular. Alterações na arquitetura e crescimento de raízes podem beneficiar ou prejudicar a captação de nutrientes pelas plantas (HERDER et al., 2010).
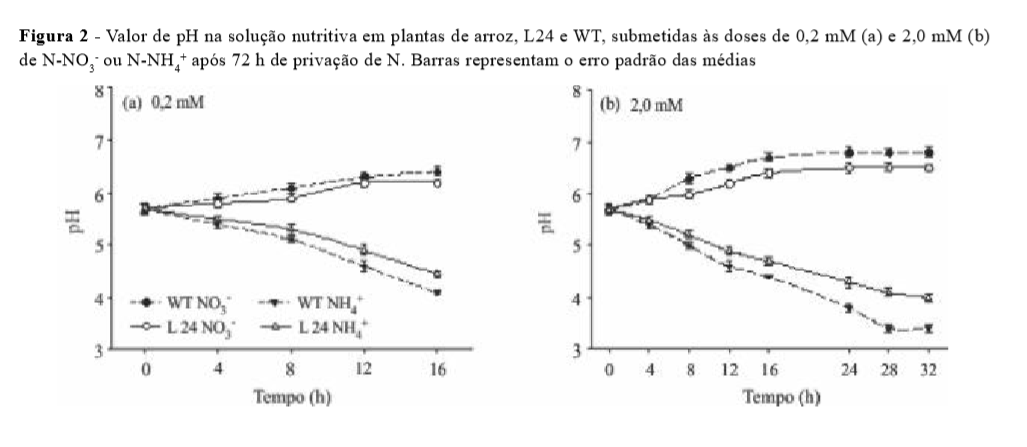
O pH da solução nutritiva foi monitorado a partir do momento em que foram aplicados os tratamentos. A L24 mostrou diminuição na capacidade de acidificação da solução quando conduzida com 0,2 e 2,0 mM de NNH4+. Também foi observada uma menor capacidade de alcalinização da solução quando as plantas foram submetidas a 0,2 e 2,0 mM de N-NO3- (Figura 2).
O fato de L24 ter mostrado baixo crescimento contribuiu para a sua menor capacidade de absorção de N (Tabela 2) e consequentemente pouca variação de pH foi observada. Entretanto, este resultado associado aos dados obtidos na cinética de absorção (Tabela 1) ratifica o menor influxo de N por L24, uma vez que para os cálculos de parâmetros cinéticos é considerada a massa de raízes
aMédias seguidas da mesma letra minúscula na linha não diferem estatisticamente pelo teste de Tukey p 0,05 de significância
das plantas, eliminando-se o efeito de diferentes massas frescas de raízes obtidas pela WT e L24.
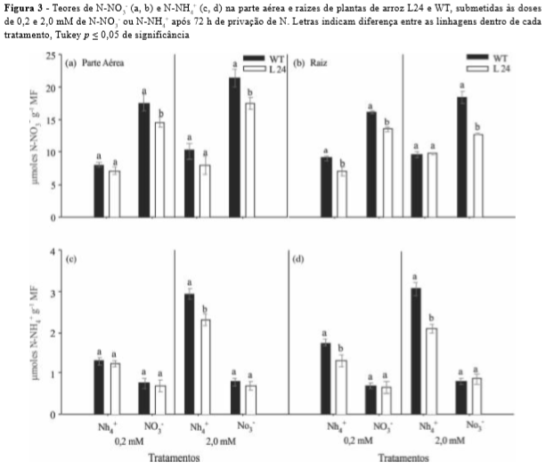
A L24 apresentou menor teor de N-NO3- em relação a WT nos tratamentos de 0,2 e 2,0 mM de N-NO3- na parte aérea e raiz (Figura 3a e b), sendo este resultado mais evidente na raiz no tratamento de 2,0 mM N-NO3-. A marcante redução no influxo deste ânion pelas plantas transformadas (Figura 1d) foi responsável pela diminuição dos teores de N-NO3- na parte aérea e raiz de L24 (Figura 3a e b). A L24 também apresentou menores teores de N-NH4+ no tratamento de 0,2 mM N-NH4+ na raiz e 2,0 mM N-NH4+ na parte aérea e raiz (Figura 3c e d). Este resultado corrobora os dados encontrados para influxo de N-NH4+ (Figura 1a e c) e sugere uma menor capacidade de redução de NO3- a NH4+ por L24 quando comparado a WT, principalmente no sistema radicular.
O NH4+ absorvido por essas plantas não deve ser acumulado no tecido vegetal, pois é responsável por sintomas de toxidez, que se expressam principalmente pelo baixo crescimento (JAMPEETONG; BRIX, 2009). Dessa forma, esse íon precisa ser assimilado em esqueletos de C e transportado na forma de aminoácidos, principalmente para a parte aérea.
Os teores de N-amino diminuíram na parte aérea e raiz em L24 (Figura 4). Foi observado que, independente do tratamento de N-NO3- ou N-NH4+ aplicado, L24 mostrou menor conteúdo de aminoácidos totais, sendo este resultado mais evidente na dose de 2,0 mM de N (Figuras 4a e b). A menor capacidade de absorção de N por L24, evidenciado pelos menores valores de influxo (Figura 1) e acúmulo de N-NO3- ou N-NH4+, resultou em menores teores de aminoácidos totais nestas plantas.
Quanto aos teores de açúcares solúveis sabe-se que nos vegetais estes podem atuar como fonte de energia prontamente disponível para o metabolismo celular e como fonte de esqueletos de C utilizados na síntese de ácidos orgânicos para assimilação do N absorvido (XU; FAN; MILLER, 2012).
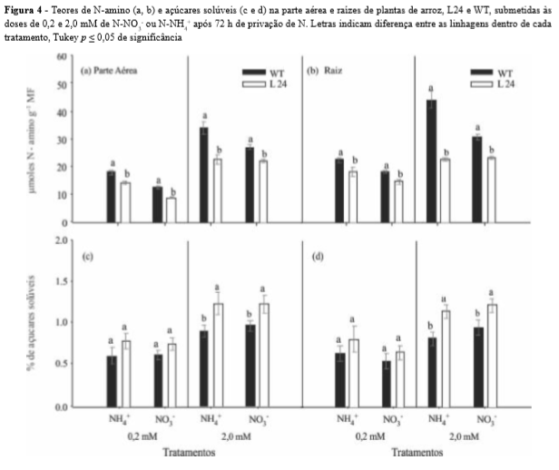
Os teores de N-amino e açúcares solúveis mostraram relação inversa, principalmente na dose de 2,0 mM de N-NO3- ou N-NH4+ em que L24 apresentou sensível aumento no teor de açúcares solúveis quando comparado a WT (Figuras 4c e d). O maior ou menor teor de N-amino e açúcares solúveis é fruto do equilíbrio existente entre esqueletos de C e N absorvido pelo vegetal sendo extensivamente relatados em trabalhos com diferentes espécies de plantas (BORGES et al., 2006; OLIVEIRA et al., 2011).
O maior teor de açúcares solúveis observado em L24 sugere uma menor capacidade destas plantas em incorporar o N absorvido em esqueletos de C, afetando Figura 4 - Teores de N-amino (a, b) e açúcares solúveis (c e d) na parte aérea e raízes de plantas de arroz, L24 e WT, submetidas às doses de 0,2 e 2,0 mM de N-NO3- ou N-NH4+ após 72 h de privação de N. Letras indicam diferença entre as linhagens dentro de cada tratamento, Tukey p 0,05 de significância assim a formação de aminoácidos essenciais para o desenvolvimento do vegetal.
A análise conjunta dos dados obtidos revelou a menor capacidade de L24 em absorver e assimilar o N disponível em solução, consequência de alterações causadas no metabolismo de C e N nas plantas transformadas. Dessa forma, os resultados mostram que o OsDof26 é possivelmente um repressor da absorção e assimilação de N pelas plantas, atuando de forma antagônica a genes que estimulam a absorção de N como o ZmDof1 (YANAGISAWA et al., 2004) e OsDof25 (SANTOS; SOUZA; FERNANDES, 2012). No entanto, ainda são necessários outros estudos que caracterizem de forma mais detalhada, os meios pelos quais a expressão deste fator de transcrição pode afetar a regulação de genes-chave envolvidos no metabolismo de C e N em plantas de arroz.
CONCLUSÕES
1.Plantas de arroz superexpressando o fator de transcrição OsDof26, L24, mostram menor massa fresca de parte aérea e raiz independente da dose e fonte de N aplicado;
2.L24 exibe parâmetros cinéticos de absorção de N desfavoráveis, com maior valor de Km e Cmín, além de menor capacidade de assimilação de N.
Fonte:
Leandro Martins Ferreira, Janiélio Gonçalves da Rocha, Leandro Azevedo Santos, Sonia Regina de Souza e Manlio Silvestre Fernandes (2015).



