Aspectos Fisiológicos do Café Conilon


1 INTRODUÇÃO
Embora exista um grande número de espécies de café, apenas Coffea arabica L. (café arábica) e Coffea canephora Pierre ex Froehner (café robusta) têm importância econômica no mercado mundial. Atualmente, cerca de 71% da produção brasileira de café é derivada de cultivares arábicas, e o restante de café robusta. A produção de café robusta, no Espírito Santo, maior produtor brasileiro, é originária exclusivamente da variedade Kouillou, ou popularmente conhecida como Conilon.
Essa variedade pertence ao grupo Guineano, apresentando grande variabilidade em relação ao porte, caules ramificados, folhas maduras com comprimento e largura menores que às das demais cultivares da espécie, folhas novas de coloração bronze, frutos vermelhos ou amarelos quando maduros e sementes de tamanhos variados (FAZUOLI, 1986).
As primeiras lavouras comerciais de café conilon foram implantadas a partir da década de 1950, com o objetivo de cultivarem-se áreas marginais ao café arábica. Em razão de sua exploração econômica ser relativamente recente, poucos estudos, ao longo do tempo, foram desenvolvidos com essa variedade e, portanto, o conhecimento sobre a fisiologia do café conilon é embrionário, se comparado com o do arábica.
Neste capítulo, procurou-se sumariar as informações existentes relativas à fisiologia do café conilon, com ênfase em aspectos que influenciam o crescimento e a produção de grãos, de forma simples, evitandose termos e conceitos específicos e detalhes mais aprofundados que, ordinariamente, interessam apenas aos fisiologistas per se. Quando pertinente, algumas informações relativas ao café arábica, especialmente sobre sistema radicular, floração e frutificação, foram extrapoladas para o café conilon.
Para uma compreensão mais detalhada da fisiologia dessa variedade, recomendam-se as revisões de DaMatta e Rena (2002), DaMatta (2004a), DaMatta e Ramalho (2006) e DaMatta et al. (2007). O leitor pode, ainda, inteirar-se de aspectos fisiológicos do café conilon sob condições de sombreamento, tratados no capítulo 19 deste livro.

2 CRESCIMENTO VEGETATIVO
Assim como ocorre na grande maioria das regiões produtoras de café do mundo, a taxa de crescimento da parte aérea do cafeeiro (crescimento dos ramos ortotrópicos e plagiotrópicos, formação de nós, expansão foliar etc.) varia sazonalmente em virtude das condições climáticas, particularmente dos regimes de chuva e de temperatura, apesar de o fotoperíodo também ter alguma influência. Dessa forma, nas regiões não equatoriais, o crescimento da parte aérea geralmente é lento no outono/inverno, durante o período seco e frio e de dias curtos, e rápido na primavera/verão, coincidindo com o início do período chuvoso e com o aumento das temperaturas e do comprimento do dia (CANNELL, 1976; AMARAL, J. A.; RENA; AMARAL, J. F., 2006).
Para o cafeeiro conilon, especificamente, há relativamente poucas informações disponíveis sobre as taxas de crescimento da parte aérea. As primeiras avaliações foram realizadas na região de Linhares, norte do Espírito Santo (SILVEIRA, 1996; SILVEIRA; CARVALHO, 1996), onde se concentra grande número de lavouras de café conilon deste Estado. Em razão da ocorrência frequente de veranicos durante a estação chuvosa e de um período seco relativamente longo, que se estende geralmente de maio a setembro, os cafezais da região norte são normalmente cultivados sob irrigação, prática que afeta, como será discutido a seguir, o crescimento da parte aérea. Nessa região, em lavouras clonais não irrigadas, a taxa de crescimento do ramo plagiotrópico é baixa nos meses de junho a setembro (0,03 cm dia-1); aumenta com o início das chuvas e atinge valores máximos em outubro (0,40 cm dia-1); reduz-se em dezembro (0,20 cm dia-1) e atinge valores mínimos em fevereiro pelas altas temperaturas; aumenta ligeiramente em março/abril (0,08 cm dia-1) e, a partir de maio, retorna aos valores mínimos (SILVEIRA, 1996; SILVEIRA; CARVALHO, 1996). Portanto, no período seco e de temperaturas mais amenas, as taxas de crescimento são baixas, enquanto no período chuvoso e de temperaturas mais elevadas, porém não extremas, as taxas de crescimento são elevadas. Com efeito, Partelli et al. (2010, 2013) observaram que, além de apresentar variações clonais, as taxas de crescimento do café conilon variaram sazonalmente, correlacionando-se fortemente com as temperaturas mínimas: apresentaram-se reduzidas sob temperaturas mínimas abaixo de 17°C (entre junho e agosto) e maiores quando as temperaturas variaram entre 17° e 31,5°C (média das temperaturas mínimas e máximas). Ademais, aqueles autores verificaram que ramos com carga pendente apresentaram menor crescimento que aqueles mais jovens. Em todo o caso, destaca-se o papel importantíssimo das primeiras chuvas após o período seco para o início do crescimento propriamente dito dos ramos plagiotrópicos. É necessário registrar, contudo, que as taxas de crescimento de ramos do café conilon ora apresentadas não são valores fixos, podendo variar não só com os clones e idade do ramo (PARTELLI et al., 2013), mas também em razão das variações temporais das condições ambientes, sobretudo as climáticas, do nível tecnológico e dos tratos culturais empregados em cada lavoura. Todavia, a variação da época de poda, seja ela realizada imediatamente após a colheita ou mais tardiamente, parece não afetar as taxas de crescimento dos ramos plagiotrópicos (MORAIS et al., 2012a).
Em lavouras irrigadas continuadamente, por sua vez, o padrão de crescimento dos ramos plagiotrópicos de plantas clonais diferencia-se consideravelmente daquele de lavouras não irrigadas. Sob irrigação, a taxa de crescimento dos ramos laterais, que é mínima em julho (0,03 cm dia-1), aumenta já em agosto (0,08 cm dia-1) e setembro (0,17 cm dia-1), diferentemente da não irrigada, e atinge taxas máximas em outubro (0,33 cm dia-1). A partir desse ponto, as taxas de crescimento assemelham-se às daquelas de lavouras não irrigadas (SILVEIRA, 1996; SILVEIRA; CARVALHO, 1996). Como será apresentado a seguir, a manutenção de uma maior área foliar em lavouras irrigadas, em comparação àquela de lavouras não irrigadas, no final do período seco (SILVEIRA; CARVALHO, 1996), poderia explicar, em parte, as diferenças nas taxas de crescimento dos ramos observadas após o período seco. Tomados em conjunto, esses dados indicam que o atraso (de aproximadamente dois meses) no restabelecimento do crescimento dos ramos plagiotrópicos em lavouras não irrigadas, comparadas às irrigadas, pode ser em parte compensado, nas últimas, por uma taxa de crescimento 20% superior em outubro, mês em que o desenvolvimento dos ramos é mais intenso (crescimento compensatório). Em outras palavras, apesar de a duração da estação de crescimento de ramos laterais nas lavouras irrigadas ser maior (julho a fevereiro) que nas não irrigadas (setembro a fevereiro, dependendo das chuvas), as taxas máximas de crescimento são observadas nas lavouras não irrigadas. Em todo o caso, esses dados permitem concluir que a disponibilidade hídrica, e não a temperatura, tem grande importância no restabelecimento do crescimento dos ramos plagiotrópicos após a estação seca. Informações dessa natureza revestem-se de grande importância, pois devem ser consideradas para a otimização do manejo das fertilizações e das podas em razão da utilização ou não da irrigação.
Em se tratando da época de formação de folhas per se, da sua taxa de expansão ao longo do ano, assim como dos fatores que a afetam, até o momento, para o cafeeiro conilon, nenhum estudo foi publicado. Todavia, a exemplo do que se verifica em café arábica, acredita-se que a produção de folhas em café conilon esteja associada intimamente com o crescimento dos caules, particularmente dos ramos plagiotrópicos, haja vista que os primórdios foliares resultam diretamente da atividade da gema apical de cada ramo lateral (RENA; MAESTRI, 1985). Portanto, a produção de folhas no café conilon, que tem relação direta com a formação de nós nos ramos laterais, provavelmente segue o mesmo padrão de crescimento supradescrito para os ramos plagiotrópicos, ou seja, é intensa na primavera/ verão e mínima no outono/inverno. Ademais, é provável, ainda, assim como ocorre no café arábica (RENA; MAESTRI, 1985), que tanto a taxa de crescimento como a área final da folha dependam da época em que se verifica a expansão, de forma que folhas formadas no início da estação chuvosa e quente apresentam maior taxa de crescimento e maior área final que folhas formadas no final da estação chuvosa, sob temperaturas mais amenas. De qualquer forma, independentemente da época de formação, a queda de folhas de plantas de café conilon intensifica-se no final da época seca, coincidindo com o período de colheita/póscolheita e de maior ação dos ventos e é afetada fortemente pela irrigação, conforme observado por Silveira (1996) e Silveira e Carvalho (1996). Esses autores verificaram reduções de aproximadamente 60% no número de folhas por nó, no período de maio a setembro, em lavouras não irrigadas e de apenas 33% durante o mesmo período, em lavouras irrigadas. Além disso, constataram maior longevidade foliar na lavoura irrigada.
O crescimento do cafeeiro conilon (planta inteira) foi avaliado por Bragança (2005), com base na produção de massa seca total ao longo de seis anos, em lavoura formada a partir da cultivar clonal ‘Emcapa 8111’ (clone 02) e conduzida em livre crescimento, em Marilândia/ES. Verificou-se que: (i) o crescimento absoluto aumentou progressivamente até atingir 15,9 kg planta-1, no 72º mês após o plantio; (ii) a taxa de crescimento absoluto aumentou, de 0,14 kg mês-1, no 3º mês, até 1,08 kg mês-1, no 48º mês, diminuindo, a seguir, até alcançar 0,46 kg mês-1, no 72º mês; (iii) a taxa total de crescimento relativo diminuiu progressivamente com a idade, variando de 0,21 kg kg-1 mês-1, no 3º mês, a 0,03 kg kg-1 mês-1, no 72º mês, provavelmente por decréscimos na taxa assimilatória líquida e na razão de área foliar; e (iv) o cafeeiro conilon apresentou padrão sigmoidal de crescimento. Esses resultados sugerem que, com o envelhecimento da lavoura, estabelece-se gradualmente um desequilíbrio entre a área foliar (associada à fotossíntese) e a massa seca total da planta que passa a ser constituída basicamente pelo excesso de ramos ortotrópicos, que se tornam fortes drenos por fotoassimilados. Como não há crescimento compensatório, principalmente em área foliar, o vigor das lavouras se reduz gradualmente e, da mesma forma, a produtividade. Tomadas em conjunto, essas informações indicam, de forma contundente, a importância e a necessidade de realização da poda na lavoura de café conilon (BRAGANÇA, 2005).
3 SISTEMA RADICULAR
Informações sobre a fisiologia do sistema radicular do cafeeiro são, sobremodo, incipientes se comparadas ao volume de informações atualmente disponível para a parte aérea. Essa escassez de pesquisa deve-se, em grande parte, às dificuldades para acessar o sistema radicular em condições naturais. Registre-se, ainda, que informações sobre o crescimento de raízes do café conilon são bem mais escassas que as disponíveis para o café arábica. De qualquer forma, recomenda-se a revisão de Rena e DaMatta (2002), na qual se discutem, de forma detalhada, vários aspectos relacionados à morfologia e ecofisiologia do sistema radicular do cafeeiro.
Apesar de existirem relatos na literatura de que a maior robustez do café robusta esteja associada à maior extensão e eficiência de seu sistema radicular, tanto na absorção de água como de nutrientes, o sistema radicular do café conilon apresenta alta plasticidade em resposta a diversos fatores. Logo, sua morfologia, arquitetura, profundidade e distribuição no solo, assim como ocorre no café arábica, pode depender da idade da planta (BRAGANÇA, 2005), do tipo de irrigação e do manejo nutricional adotados (BARRETO et al., 2006), da relação entre fonte e dreno (ALVES et al., 2011), do método de formação de mudas, i.e., se propagadas vegetativamente ou a partir de sementes (JESUS; CARVALHO; SOARES, 2006; PARTELLI et al., 2006, 2014b; SILVA et al., 2010), das características químicas do solo (MOTA et al., 2006), da calagem em subsuperfície (RODRIGUES et al., 2001), das práticas culturais (RENA; GUIMARÃES, 2000; RENA; DaMATTA, 2002), da competição e das espécies de plantas daninhas (RONCHI; TERRA; SILVA, 2007), da densidade de plantio (CASSIDY; KUMAR, 1984; RONCHI et al., 2015b); e do material genético (ALFONSI et al., 2005; BRAGANÇA, 2005; PINHEIRO et al., 2005; SILVA et al., 2010; COVRE et al.,2013; RONCHI et al., 2015b).
Trabalhos recentes, realizados ao longo de vários anos no Município de Vila Valério, região norte do Espírito Santo produtora de café conilon, apontaram que, apesar de a produtividade de plantas propagadas por estacas ser maior que aquelas propagadas por sementes, ambas as formas de propagação originaram sistemas radiculares semelhantes (PARTELLI et al., 2006, 2014b). Portanto, o sistema radicular do café conilon, principalmente as raízes absorventes (menores que 1,0 mm de diâmetro), avaliado por meio da área, comprimento e volume de raízes por volume de solo, seja em plantas oriundas de propagação vegetativa ou seminífera, concentra-se na camada superficial do solo: de 40% a 50% até 0,1 m e de 60% a 65% até 0,2 m de profundidade (Figuras 1 e 2), com padrão marcante de redução, à medida que se aprofunda no perfil do solo (PARTELLI et al., 2006; 2014b). Essa redução é explicada, parcialmente, pela menor concentração de alguns macronutrientes em profundidade (PARTELLI et al., 2014b). Em todo o caso, na principal região produtora de café conilon do norte do Estado do Espírito Santo, em que predomina solo do tipo Latossolo Vermelho Amarelo distrófico (LVd11), com presença de camada adensada subsuperficialmente, o sistema radicular pode concentrar-se na camada de 0-0,45 m de profundidade, apesar de uma pequena quantidade de raízes conseguir penetrar a camada adensada (SILVA et al., dados não publicados) (Figura 1).

De certa forma, a estrutura e a distribuição das raízes do café conilon são semelhantes àquelas do café arábica (RENA, 1998; RENA; DaMATTA, 2002; RONCHI et al., 2015b). Entretanto, ao se avaliar o sistema radicular de uma lavoura de conilon, cultivada no espaçamento de 2,0 x 1,0 m, Partelli et al. (2014b) constataram que o comprimento e o volume de raízes por unidade de solo foram maiores na região do solo, sob a copa, voltada para a entrelinha (onde as adubações de solo eram feitas), em comparação aos valores medidos entre plantas na linha. De forma contrária, em café arábica, Ronchi et al. (2015b) mostraram que a 0,25 m do tronco e, sobretudo, entre plantas adjacentes na linha de plantio, para diferentes espaçamentos de cultivo, é a região do solo onde se concentra o sistema radicular do cafeeiro (terceira safra), provavelmente, em decorrência da presença do sistema de fertirrigação nessa posição. De qualquer forma, a distância efetiva do sistema radicular de C. canephora cv. Apoatã não ultrapassa 0,8 m do tronco, no sentido da entrelinha (BARRETO et al., 2006).
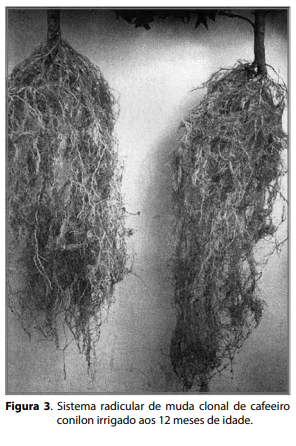
Mesmo diante do fato de que a irrigação permite a formação de mudas clonais com um sistema radicular mais vigoroso (Figura 3), é preciso considerar que a irrigação afeta o padrão de crescimento radicular, reduzindo a profundidade de penetração da raiz pseudopivotante e estimulando o desenvolvimento de raízes primárias e secundárias nas camadas mais superficiais do solo (RENA, 1998). Com efeito, em lavouras adultas de diferentes cultivares de café arábica, cultivadas sob fertirrigação no cerrado mineiro, a abundância de raízes por volume de solo é maior a 0,1 m que a 0,2 m ou 0,4 m de profundidade para vários arranjos de plantas na linha de cultivo (RONCHI et al., 2015b). Não obstante, esses mesmos autores verificaram que a redução no espaçamento, entre plantas, na linha promoveu aumento na massa de matéria seca, comprimento, volume e área superficial total de raízes por volume de solo, sem alterar o comprimento e a superfície específica de raízes e sem aprofundar o sistema radicular. Assim, os resultados obtidos em café arábica por Ronchi et al. (2015b) corroboram e sustentam os argumentos de Guarçoni M. (2011), de que o aumento na densidade de plantas em lavouras de café conilon não precisa necessariamente ser acompanhado de uma elevação proporcional na aplicação de fertilizantes por área. Isso porque a maior abundância ou distribuição quantitativa de raízes por volume de solo, em cafeeiros adensados, pode compensar um menor input de fertilizantes. Tomados em conjunto, esses resultados podem explicar, pelo menos em parte, as altas produtividades obtidas em lavouras de café conilon cultivadas em sistemas mais adensados, conforme se discute no capítulo 11 deste livro.
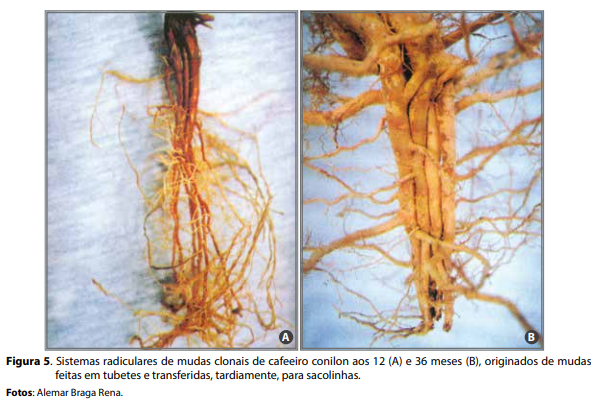
Outro fator que pode afetar a qualidade do sistema radicular do café conilon e que, no passado, constituiu-se em grandes problemas à cafeicultura Figura 2. Sistema radicular do cafeeiro conilon (clone 02) em lavoura adulta, na Fazenda Experimental de Marilândia/ES. Fotos: José Antônio Lani. Figura 3. Sistema radicular de muda clonal de cafeeiro conilon irrigado aos 12 meses de idade. capixaba, é a formação de mudas a partir de estacas (clones) mantidas por longos períodos em tubetes. Nesse caso, verificam-se raízes mal-formadas e raquíticas (Figura 4). Essas mudas quando levadas ao campo, apresentam-se com raízes enoveladas, superficiais e deformadas (Figura 5). No longo prazo, observam-se redução da absorção de água e de nutrientes e morte de plantas, principalmente em anos de carga intensa (RENA, 1998).

O volume, a área superficial, o comprimento e a quantidade de matéria seca de raízes de café conilon podem variar conforme o clone utilizado. Nesse contexto, Pinheiro et al. (2005) verificaram que a produção de massa seca de raízes por planta de café conilon com dez meses de idade, cultivada sem restrição hídrica, em vasos de 120 L, foi de 0,176; 0,187; 0,351 e 0,268 kg para os clones 14, 46, 109A e 120, respectivamente. É importante ressaltar que maior quantidade de massa seca de raízes não está, necessariamente, associada a maiores profundidades do sistema radicular. Por exemplo, nesse trabalho de Pinheiro et al. (2005), a profundidade média foi de 0,75 m nos clones 14 e 120, e de aproximadamente 0,50 m nos clones 46 e 109A. Esses resultados foram confirmados posteriormente por Silva et al. (2010) para os clones 120 e 109A, também cultivados em vasos. Esse conhecimento sobre o sistema radicular é importante, pois numa situação de deficit hídrico, o retardamento da desidratação, associado à profundidade, e não à matéria seca per se do sistema radicular, pode garantir maior tolerância à seca a alguns materiais genéticos.

Em mudas clonais formadas em viveiro, Covre et al. (2013) constataram que todos os clones que compõem a cultivar de café conilon ‘Vitória Incaper 8142’ apresentaram sistemas radiculares semelhantes quando eles foram avaliados pelo diâmetro médio ponderado e pelo volume de raízes. Todavia, os clones V3, V8 e V10 destacaramse dentre os demais quanto à área superficial de raízes. Constata-se, portanto, que variações morfológicas dos sistemas radiculares de diferentes clones de café conilon podem ou não ocorrer dependendo da variável utilizada para sua quantificação. Isso também ocorre para o café arábica, assim como marcante variação entre cultivares (RONCHI et al., 2015b).
Bragança (2005) demonstrou em condições de campo que o sistema radicular do café conilon varia fortemente com sua idade. Nesse caso, a produção de biomassa seca do sistema radicular do cafeeiro conilon (clone 02) foi avaliada ao longo de 72 meses, após o transplantio, apresentando crescimento segundo o modelo logístico Y = 2,87/(1+26,13 e (-0,0835 t)), de forma que aos 12, 36 e 72 meses (t), o cafeeiro apresentou 0,27; 1,25 e 2,69 kg planta-1 de massa seca de raízes, respectivamente. Além desses resultados, Bragança (2005) verificou também que a taxa de crescimento absoluto do sistema radicular aumentou de 0,03 kg mês-1, no terceiro mês, até 0,18 kg mês-1 no 40º mês, diminuindo a seguir até alcançar 0,05 kg mês-1 no 72º mês, e que a taxa de crescimento relativo reduziu de 0,21 kg kg-1 mês-1, no terceiro e sexto mês, a 0,02 kg kg mês-1 no 72º mês, após o transplantio.
O sistema radicular do cafeeiro conilon também pode ser afetado pela carga pendente. Com efeito, em lavoura irrigada de café conilon (clone 02 - de maturação precoce), conduzida em livre crescimento, Bragança (2005) verificou que, num ano de superprodução (quarta safra – 200 sc. benef./ha, diferentemente dos anos anteriores, nos quais a produção foi regular e bem inferior), o sistema radicular do cafeeiro reduziu-se de 3,30 kg planta-1, em outubro, para 1,50 e 1,60 kg planta-1 em janeiro e abril, respectivamente. Nesse caso, o crescimento foi retomado durante o inverno, de forma que, em julho e outubro seguintes, aqueles valores atingiram 2,18 e 3,5 kg planta-1, respectivamente. Esses resultados reforçam as hipóteses de que, em anos de carga elevada, há o esgotamento da planta e o consequente comprometimento (morte) do sistema radicular, e de que há crescimento deste último durante o inverno, principalmente em lavoura irrigada. Em suma, em contraste ao observado na parte aérea, nos períodos secos e frios (inverno) ou secos e quentes (veranicos), acredita-se que o crescimento das raízes, principalmente daquelas de menor diâmetro e mais profundas, não cessa, uma vez que nas proximidades dessas raízes a temperatura do solo dificilmente atingirá níveis extremos (conquanto haja água disponível), se comparados à amplitude térmica a que a parte aérea está exposta. Portanto, a redução do crescimento da parte aérea parece preceder o crescimento do sistema radicular (RENA, 1998).
4 CRESCIMENTO REPRODUTIVO
4.1 FLORAÇÃO
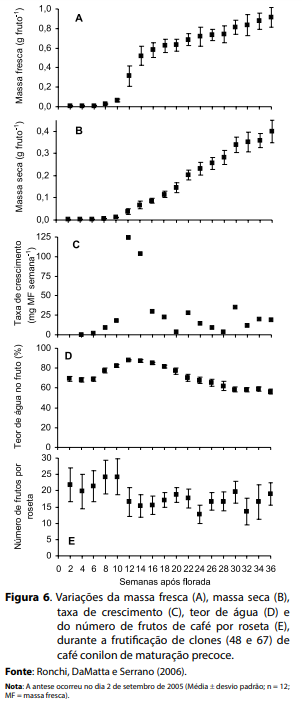
O café é uma espécie tropical de floração gregária, ou seja, todas as plantas individuais de uma certa extensão geográfica florescem simultaneamente (RENA; BARROS, 2004). Todavia, um número variável de floradas com intensidades e temporalidades variáveis são, geralmente, observadas a campo. Em café conilon, as inflorescências (glomérulos) são formadas a partir de gemas seriadas (sendo um glomérulo por cada gema) localizadas aleatoriamente nas axilas das folhas de ramos laterais que se formaram na estação de crescimento do ano corrente, de forma que a floração depende estreitamente do crescimento dos ramos plagiotrópicos (MOENS, 1968, apud BARROS; MAESTRI; COONS, 1978). Silveira e Carvalho (1996) observaram, em café conilon, 3,3 glomérulos por axila foliar e 3,4 flores por glomérulo, em média. Portanto, apenas uma roseta de um dado ramo plagiotrópico produziria, potencialmente, de 22 a 24 chumbinhos. Avaliações conduzidas por Ronchi, DaMatta e Serrano (2006) em clones de café conilon de maturação precoce dos frutos parecem suportar essas considerações: imediatamente após a florada, no início da fase de chumbinho e, independentemente de irrigação, cada roseta localizada na porção mediana de ramos plagiotrópicos do terço superior da planta continha, de fato, uma média de 24 chumbinhos (Figura 6E).
Erroneamente, o termo floração muitas vezes é usado para designar apenas a abertura das flores de café (antese). Entretanto, a antese representa apenas uma (a última) das várias fases da floração. Segundo Rena e Barros (2004), esse processo complexo pode ser dividido em quatro fases: (i) iniciação; (ii) diferenciação; (iii) período de dormência do botão floral; e (iv) abertura da flor ou florada. Após as duas primeiras fases, a gema seriada, que poderia dar origem a um ramo ou a uma inflorescência, passa de vegetativa a reprodutiva, levando, irreversivelmente, à formação da inflorescência (RENA; MAESTRI, 1985; RENA; BARROS; MAESTRI, 2001; RENA; BARROS, 2004). Para o café conilon, particularmente, os eventos associados às fases iniciais da floração são totalmente desconhecidos. Na literatura, há menção a apenas um trabalho, realizado há aproximadamente seis décadas, em que se estudou a morfogênese ou morfologia de gemas de C. canephora cv. Robusta (MOENS, 1968, apud BARROS; MAESTRI; COONS, 1978). Verificou-se que o primórdio floral com altura inferior a 0,3 mm ainda não havia sofrido diferenciação, que ocorreu somente quando o ápice cônico e proeminente do meristema tornou-se largo e achatado. Externamente, a olho nu, a constatação de estípulas completamente comprimidas ao ramo lateral e de que o nó não mostra qualquer alargamento acima delas indica que a gema floral ainda não está diferenciada floralmente (MOENS, 1968, apud BARROS; MAESTRI; COONS, 1978).
Os botões florais, após passarem por um período de dormência, tornam-se sensíveis ou fisiologicamente maduros e reagem aos estímulos externos desencadeadores de um rápido crescimento, que dura cerca de dez dias, levando à abertura das flores. Em condições naturais, a florada em café é provocada pelas primeiras chuvas da estação após um período de seca (BARROS; MAESTRI; COONS, 1978; RENA; BARROS; MAESTRI, 2001), que ocorre, geralmente, em lavouras de conilon não irrigadas do Espírito Santo, em agosto/setembro. É importante ressaltar, entretanto, que informações acerca dos fatores luz, temperatura, disponibilidade hídrica, chuvas, relação carbono/nitrogênio, níveis de carboidratos, carga de frutos e hormônios, que supostamente regulam as diferentes fases da floração, são praticamente desconhecidos em café conilon, assim como os eventos de natureza bioquímica, fisiológica e morfológica, que ocorrem na gema durante sua diferenciação, dormência e nos momentos que antecedem a florada. Mesmo para o arábica, o processo de floração não é conhecido em profundidade (BARROS; MAESTRI; COONS, 1978; RENA; BARROS; MAESTRI, 2001; RENA; BARROS, 2004; DaMATTA; CAVATTE; MARTINS, 2012).
O desenvolvimento do botão floral em café conilon, do período final de dormência até a antese, foi estudado em Linhares, em lavouras conduzidas com e sem irrigação apesar de as avaliações terem sido iniciadas tardiamente (segunda quinzena de junho), poucas semanas antes da antese, quando os botões florais estavam totalmente diferenciados, porém bem visíveis a olho nu (SILVEIRA; CARVALHO, 1996). Nesse caso, foram definidos quatro estádios de desenvolvimento de botões florais: achatado, ovalado de coloração verde, ovalado de coloração verde-clara e flor propriamente dita. Além disso, o número de botões por nó, em cada estádio, foi avaliado em várias épocas. Verificouse que a irrigação atrasou o desenvolvimento do botão floral em relação àquele observado em lavouras não irrigadas: (i) aos 60 dias antes da florada, as plantas não irrigadas apresentavam a mesma porcentagem (~50%) de botões florais no estádio achatado e ovalado de coloração verde, ao passo que a porcentagem de cada um desses estádios nas plantas irrigadas era de 66% e 33%, respectivamente; (ii) aos 30 dias antes da florada, nas plantas sem irrigação, os únicos estádios de botões florais presentes nos nós eram praticamente flores (poucas) e, predominantemente, botões ovalados de coloração verde-clara; todavia, nas plantas irrigadas, nessa mesma época, havia aproximadamente 5% de botões achatados, 25% de botões ovalados de coloração verde e 60% de botões de coloração verde-clara; (iii) durante a florada, verificaram-se apenas flores nos nós das plantas não irrigadas, enquanto nas irrigadas, além de flores, 20% dos botões florais ainda estavam dormentes. Consequentemente, Silveira e Carvalho (1996) constataram que, apesar do maior número de glomérulos por axila foliar, as plantas irrigadas apresentaram menor número de flores por glomérulo que as plantas não irrigadas. Tomados em conjunto, esses dados sugerem, a exemplo do que ocorre com o café arábica (BARROS; MAESTRI; COONS, 1978; RENA; BARROS; MAESTRI, 2001; DaMATTA et al., 2007), que um período seco pode contribuir para a maturação mais concentrada e antecipada dos botões florais de café conilon, com a concentração, no tempo, da florada.
4.1.1 Concentração da florada pela imposição de deficit hídrico
Um problema importante relacionado à produção de café conilon (assim como na de café arábica) refere-se à grande variação nas porcentagens de grãos cerejas, verdes, passas ou secos, comumente verificada por ocasião da colheita (THOMAZINI et al., 2011). Essa desuniformidade de maturação dos frutos, além de causar inúmeros prejuízos diretos ou indiretos (RENA; BARROS, 2004; GUERRA et al., 2006; DaMATTA et al., 2007), compromete a qualidade final do café, sobretudo quando a porcentagem de grãos verdes aumenta e a de cereja diminui (THOMAZINI et al., 2011). Além disso, a colheita de grãos verdes, ainda não granados, pode acarretar quedas significativas de renda do café (perda de peso) (MATIELLO, 1998; SOUZA; SANTOS; VENEZIANO, 2005; MATIELLO et al., 2010).
Nas regiões não equatoriais em que comumente o cafeeiro conilon é cultivado, como, por exemplo, no norte do Estado do Espírito Santo e sul da Bahia, a desuniformidade de maturação dos frutos é praticamente inevitável, uma vez que ocorrem várias floradas no período de julho a novembro. Isso se dá por dois fatores principais (DaMATTA et al., 2007): i) a iniciação e diferenciação de cada gema floral, que são as primeiras fases do processo de floração do cafeeiro (verificadas no final do verão/início do outono), não ocorrem no mesmo momento. Consequentemente, nas fases finais da floração, as gemas ficarão maduras ou aptas para responder ao estímulo de abertura da flor em momentos diferentes; ii) a ocorrência de chuvas esporádicas, muitas vezes de baixa intensidade, durante as fases finais do desenvolvimento do botão floral (geralmente em agosto), leva à abertura daquelas poucas gemas maduras, geralmente originando floradas fracas e esparsas.
Os fatores (genético e climático) anteriormente discutidos são de difícil controle, particularmente nas lavouras não irrigadas. Todavia, nas lavouras irrigadas, sobretudo naquelas regiões em que predomina uma estação seca bem definida, entre maio e setembro (como comumente ocorre na principal região produtora de café conilon brasileira, a região norte capixaba), o manejo da irrigação durante o período pré-florada pode ser a chave para obterem-se floradas concentradas. A aplicação de deficit hídrico controlado pela suspensão da irrigação, num período específico que antecede a abertura das flores, contribui para a concentração da florada. Isso está associado ao fato de que o deficit hídrico permite que maior número de gemas florais (aquelas mais atrasadas) complete os eventos morfológicos e fisiológicos necessários à sua completa maturação. Logo, mais gemas em toda a planta estarão aptas a se abrir de uma única vez, em resposta à chuva ou ao retorno da irrigação, levando ao que popularmente se denomina de concentração da florada no tempo (DaMATTA et al., 2007).
É possível, portanto, conseguir maior porcentagem de grãos cerejas em lavouras submetidas ao deficit hídrico, em comparação à daquelas irrigadas continuadamente (MATIELLO et al., 2010; RONCHI et al., 2015a). Infelizmente, a implementação do deficit hídrico adequado, em condições de campo, é tarefa difícil, de forma que a definição da época ou momento adequado para a sua imposição é um dos aspectos de maior controvérsia acerca da concentração de florada no Brasil (SOARES et al., 2005). De qualquer forma, estabelecer datas fixas do calendário para suspender e retomar a irrigação, sem aferir o nível de deficit e o estádio do botão floral, como proposto por Guerra, Rocha e Rodrigues (2005) para o café arábica, carece de maior comprovação científica (RONCHI et al., 2015a).
Apesar desse método (uso de datas fixas) ser de fácil implantação pelos produtores, no que concerne ao manejo da irrigação, mesmo para as regiões com estação seca bem definida, diversos fatores como características do solo, temperatura e umidade relativa do ar e suas variações diárias, deficit de pressão de vapor, índice de área foliar do cafeeiro, espaçamento e orientação de plantio, idade da planta e arquitetura de copa, clones e cultivares, profundidade do sistema radicular, entre outras, afetam a taxa de perda d’água pelas plantas (DaMATTA et al., 2007; SILVA et al., 2009) e, consequentemente, o tempo necessário para se atingir o nível de deficit desejado e adequado à abertura floral para uma dada lavoura (SOARES et al., 2005; RONCHI et al., 2015a).
Para fundamentação teórica mais detalhada acerca dos aspectos fisiológicos relacionados à aplicação do deficit hídrico para concentração da florada, recomenda-se a leitura de DaMatta et al. (2007) e Ronchi et al. (2015a). Ressalta-se, entretanto, que quase a totalidade de trabalhos publicados na última década sobre esse tema (ver referências em Ronchi et al., 2015a) refere-se à espécie C. arabica, e os resultados descritos são muito variáveis em função da diversidade de condições experimentais de cada trabalho. Para o conilon, apenas poucos trabalhos de cunho científico foram recentemente realizados, todos na região norte do Estado do Espírito Santo, cujos principais resultados são descritos a seguir.
No Município de Jaguaré/ES, Marsetti et al. (2013) avaliaram os efeitos de diferentes períodos de suspensão da irrigação (por aspersão) na uniformidade da florada do clone 02, com cinco anos de idade. Além do tratamento irrigado ininterruptamente, aqueles autores testaram a suspensão da irrigação por 3, 5, 7, 9 ou 12,5 semanas, a partir de 5 de julho, sendo a contagem de flores realizada no décimo dia após o fim de cada período, ou seja, dez dias após o retorno da irrigação. Constataram, então, que a porcentagem de flores (ou intensidade da florada) foi de 14%, 14%, 57%, 96% e 97%, respectivamente, para aqueles tratamentos com períodos de suspensão da irrigação, sem observarem qualquer efeito destes sobre o crescimento dos ramos plagiotrópicos. Isso indica, portanto, um efeito positivo da interrupção da irrigação na concentração da florada do conilon. Apesar de não terem quantificado o nível de deficit hídrico atingido ao final de cada período, o que seria primordial para a explicação dos resultados (RENA; BARROS, 2004; RONCHI et al., 2015a), Marsetti et al. (2013) concluíram que a baixa porcentagem de flores verificadas após três, cinco ou mesmo sete semanas de suspensão da irrigação ocorreu, pois o deficit hídrico foi aplicado em épocas inadequadas, quando a maioria dos botões florais ainda não estavam fisiologicamente maduros. E isso é uma das principais causas do insucesso do uso do deficit hídrico para concentração da florada do cafeeiro, como constatado para o café arábica (CRISOSTO; GRANTZ; MEINZER, 1992; SOARES et al., 2005; BOMFIM NETO, 2007; SILVA et al., 2009; RONCHI et al., 2015a).
Um resultado relevante encontrado por Marsetti et al. (2013) para o conilon, porém não discutido pelos autores, foi o fato de que o tratamento sob irrigação contínua também proporcionou elevada porcentagem de flores (94%), semelhante à daquela de plantas sob deficit hídrico por mais de sete semanas. Provavelmente, essa florada originou-se após uma chuva ocorrida em fins de setembro, que quebrou a dormência dos botões florais fisiologicamente maduros, mas mantidos sob elevado potencial hídrico, pela alteração repentina nos componentes climáticos atmosféricos levandoos à antese, como também verificado e discutido recentemente para o café arábica (RONCHI et al., 2015a).
Em São Mateus/ES, Souza et al. (2014) avaliaram a uniformidade de maturação de frutos de nove clones da cultivar Vitória Incaper 8141 em resposta à suspensão da irrigação no período de 16/06 a 15/08 (60 dias), portanto, na pré-florada. Apesar de que aparentemente essa época não tenha sido a mais adequada para a imposição do deficit hídrico, devido à precocidade do desenvolvimento do botão floral, conforme discutido anteriormente, para Marsetti et al. (2013), a porcentagem média de cerejas, considerando todos os clones avaliados, aumentou significativamente de 71%, no tratamento sob irrigação contínua, para 76% naqueles tratamentos com aplicação de deficit hídrico. Entretanto, essa resposta positiva de aumento da porcentagem de cerejas, em detrimento aos outros estádios avaliados (verdes e secos), foi verificada apenas em seis dos nove clones avaliados. Ademais, a intensidade da resposta variou levemente entre esses seis clones. Dessa forma, além de se caracterizar um efeito discreto do deficit hídrico sobre a uniformidade de maturação dos frutos do café conilon, pode-se sugerir que essa resposta exibe variações clonais, o que é explicável considerando-se que clones de conilon atingem diferentes níveis de deficit hídrico após um mesmo período de suspensão da irrigação, em decorrência das suas diferentes taxas de perda d´água (PINHEIRO et al., 2005; SILVA et al., 2013). Além disso, há o fato de que a magnitude do deficit hídrico é fator determinante para a quebra da dormência dos botões florais maduros (RONCHI et al., 2015a).
Considerando em conjunto as informações apresentadas até aqui, há praticamente um consenso para o arábica e, agora, para o conilon de que o estádio de desenvolvimento do botão floral e o nível de deficit hídrico do cafeeiro, ambos associados às condições climáticas predominantes nas fases finais da floração, sejam os principais critérios a serem considerados para suspensão e retomada da irrigação, visando ao sucesso na concentração da florada. A ocorrência de uma estação seca bem definida favorece a aplicação do deficit hídrico nas lavouras irrigadas, garantindo floradas e maturação dos frutos mais uniformes e, consequentemente, melhoria na qualidade do café (SOUZA et al., 2014). Não obstante, outras vantagens secundárias da interrupção da irrigação são a economia de água, energia elétrica e mesmo de mão de obra (MARSETTI et al., 2013), podendo, portanto, reduzir significativamente o custo de produção do conilon.
Apesar dessas poucas evidências científicas, conclui-se que a técnica para concentração de florada do conilon pela sua exposição ao deficit hídrico é perfeitamente viável, todavia, não existe recomendação padrão ou receita acabada. Assim, faz-se mister monitorar o clima e fazer pesquisas aplicadas em cada situação para se definir, com precisão local, o melhor momento de aplicação do deficit hídrico (e sua magnitude), sem comprometer o crescimento, a produtividade e a longevidade da lavoura de conilon.
4.2 FRUTIFICAÇÃO
Após a abertura das flores, inicia-se a fase de frutificação do cafeeiro, na qual ocorre o pegamento, o desenvolvimento e a maturação dos frutos. O pegamento final de frutos de café conilon, avaliado como a percentagem de frutos de café presentes nos ramos após a florada em relação ao número inicial de flores, foi de aproximadamente 30% para plantas não irrigadas ou irrigadas continuamente, ao passo que a maior taxa de queda de frutos ocorreu no final da fase chumbinho e, principalmente, durante a fase de expansão rápida dos frutos (SILVEIRA; CARVALHO, 1996). De forma semelhante, verificou-se, em café conilon, que o número médio inicial de 24 chumbinhos por roseta reduziu-se para aproximadamente 15 frutos-cereja por roseta, evidenciando uma queda de 38% de frutos nesse período (Figura 6) (RONCHI; DaMATTA; SERRANO, 2006). Além disso, essa queda ocorreu preponderantemente durante a fase inicial da expansão rápida dos frutos, cujas causas prováveis são citadas e/ou discutidas por Rena, Barros e Maestri (2001) e Ronchi et al. (2005).
O período de crescimento ou desenvolvimento dos frutos é geralmente dividido, em café arábica, em cinco fases: fase 1 - chumbinho; fase 2 - expansão rápida; fase 3 - formação do endosperma; fase 4 - endurecimento do endosperma (granação); e fase 5 - maturação dos frutos. Essa divisão também pode ser aplicada ao café conilon. O padrão de crescimento segue, na maioria das vezes, um modelo sigmoidal duplo (LEON; FOURNIER, 1962; SALAZAR-GUTIÉRREZ et al., 1994), mas pode variar, dependendo das condições climáticas e da altitude (LAVIOLA et al., 2007), das variáveis utilizadas para medir o crescimento do fruto (e.g., comprimento, diâmetro, volume, massas seca e fresca), da frequência de medições, dos modelos estatísticos e das espécies (WORMER, 1964; SALAZAR-GUTIÉRREZ et al., 1994). Em café robusta, por exemplo, existem divergências quanto ao tipo de curva utilizado para caracterizar o crescimento de frutos nos poucos trabalhos publicados com essa espécie, no mundo (DANCER, 1964; RAMALAH; VASUDEVA, 1969; OYEBADE, 1976; SRINIVASAN; RAJU; VISHVESHWARA, 1978), apesar de Rena, Barros e Maestri (2001) sugerirem que o padrão sigmoidal seja comum a ambas as espécies, café arábica e robusta. Especificamente para o café conilon, Silveira (1996) verificou que o crescimento de frutos, com base no acúmulo de massa seca, seguiu o modelo sigmoidal duplo, quando cultivado sem irrigação, e o sigmoidal simples, quando irrigado. Recentemente, Partelli et al. (2014c) encontraram um padrão sigmoidal simples para o acúmulo de matéria seca de frutos de café conilon de maturação precoce (clone 12 V), intermediária (clone 10 V) e tardia (clone 13 V). Ademais, de forma geral, pode-se afirmar que a curva de acúmulo de macronutrientes pelos frutos desses clones seguiu um padrão similar ao daquele da matéria seca.
Atualmente, a grande maioria das lavouras de café conilon é formada por cultivares clonais, as quais são compostas por um determinado número de clones. Algumas dessas cultivares distinguem-se entre si fenotipicamente, basicamente, pela época de maturação dos frutos, ou seja, os frutos de todos os clones de uma determinada cultivar completam, simultaneamente, sua maturação, que pode ser precoce, intermediária ou tardia (BRAGANÇA et al.,1993). Portanto, em relação à maturação, existem clones chamados precoces, intermediários e tardios. É importante ressaltar, entretanto, que a florada desses clones ocorre na mesma época, em agosto/ setembro, por ocasião das “chuvas de florada”. Contudo, o período posterior, da abertura da flor à completa maturação (e colheita), é diferenciado para cada clone: clone precoce – 34 semanas (colheita em maio), clone intermediário – 41 semanas (colheita em junho), clone tardio – 45 semanas (colheita em julho) (BRAGANÇA et al., 1993, 2001). Resultados recentes também comprovaram essa variação entre clones quanto ao período necessário para a completa maturação do fruto, a qual ocorre aos 216, 244, 244 e 300 dias após a antese para clones de maturação precoce, intermediária, tardia e supertardia respectivamente (PARTELLI et al., 2014c). Mesmo apresentando idêntico padrão (sigmoidal) de crescimento, a duração de cada fase é variável de acordo com o clone analisado (PARTELLI et al., 2014c). Ressalte-se que a duração de cada fase e, consequentemente, o período total necessário para a completa maturação dos frutos dependem não apenas do material genético, mas da forte interação entre fatores como altitude, temperatura, disponibilidade hídrica e soma térmica (ou grausdia) (LAVIOLA et al., 2007; PETEK; SERA; FONSECA, 2009).
Estudos sobre o padrão de crescimento dos frutos revestem-se de grande importância científica e agronômica, pois a partir das curvas de crescimento é possível inferir as fases em que a demanda do fruto (e da planta) por água e nutrientes é mais ou menos pronunciada. Podendo-se, portanto, otimizar o manejo de água e de fertilizantes e, consequentemente, reduzir custos e aumentar tanto a produtividade da lavoura como a qualidade dos frutos. Nesse sentido, trabalho realizado em Sooretama/ES buscou determinar o padrão de crescimento de frutos de diversos clones de café conilon de maturação precoce, intermediária e tardia, cultivados com e sem irrigação (RONCHI; DaMATTA; SERRANO, 2006).
Considerando que o crescimento do fruto do café conilon se dá em cinco fases distintas, semelhantemente às daquelas para o café arábica, para cafeeiros de maturação precoce, elas teriam a seguinte duração (Figuras 6 e 7): fase 1 - estádio de chumbinho: da florada até a sexta semana seguinte duração de oito semanas; fase 2 - crescimento rápido: da sexta à 16ª semana após a florada – duração de dez semanas; fase 3 - crescimento suspenso: da 16ª à 18ª semana após a florada – duração de duas semanas; fase 4 - granação: da 18ª à 28ª semana após a florada – duração de dez semanas; e fase 5 - maturação: da 28ª à 36ª semana após a florada – duração de oito semanas (RONCHI; DaMATTA; SERRANO, 2006). É importante ressaltar, contudo, que a transição entre fases é um processo contínuo (não abrupto) e, portanto, os limites inferiores e superiores ora definidos para cada fase são valores aproximados.
Durante a fase de chumbinho, não houve acúmulo de massa fresca (Figura 6A) e massa seca (Figura 6B) e, portanto, a taxa de crescimento foi praticamente nula (Figura 6C). Nessa fase, o teor de água do fruto (~86,5%; Figura 6D) e o número de frutos por roseta (Figura 6E) permaneceram inalterados. Ressalte-se que os frutos nessa fase não podem ser considerados dormentes, pois apresentam elevada taxa de respiração (CANNELL, 1976).
Durante a fase de expansão rápida, a massa fresca do fruto aumentou em aproximadamente 58 vezes (de 0,010 g para 0,580 g; Figura 6A), e a sua massa seca em 25 vezes (de 0,003 g para 0,075 g; Figura 6B), de forma que a taxa de crescimento, que era de 1,8 mg (massa fresca) por semana, no final da fase chumbinho, atingiu valores máximos de 124 mg (massa fresca) por semana (Figura 6C). Tanto o aumento na quantidade de massa seca como na quantidade de água contribuíram simultaneamente para explicar o aumento exponencial verificado na massa fresca do fruto. Todavia, a quantidade de água aumentou, proporcionalmente, mais que o aumento da massa seca, de forma que, durante a fase de expansão rápida, o teor de água no fruto atingiu seus valores máximos, superiores a 88% (Figura 6D). Nessa fase, provavelmente pela competição entre frutos por espaço na axila da folha, verificou-se a sua intensa queda chegando, em média, a dez frutos por roseta (Figura 6E).
Durante a fase 3, ainda houve um pequeno aumento na massa fresca e na massa seca do fruto; entretanto, ocorreu queda acentuada na taxa de crescimento relativa e redução discreta no teor de água no fruto (Figura 6). Acredita-se, a exemplo do que se verifica em café arábica (WORMER, 1964), que tanto o pergaminho como o fruto per se atinjam seus tamanhos máximos nessa fase.
Durante a fase 4 ou período de enchimento do endosperma – granação (de meados de dezembro a meados de março), enquanto a massa fresca aumentou discretamente (18,5%), a massa seca do fruto teve significativa elevação de 143% (Figura 6C). Por causa desse aumento, o teor de água do fruto caiu para valores inferiores àqueles da fase chumbinho, ou seja, para 62% (Figura 6D).
Ao final da granação, a semente atinge sua maturidade fisiológica, quando sua massa seca máxima é atingida (ASTOLF et al., 1981). A partir de então, inicia-se a quinta e última fase do desenvolvimento do fruto, a maturação, que envolve, predominantemente, alterações no pericarpo (casca do fruto), cujo volume aumenta (RENA; MAESTRI, 1985). Isso pode explicar o pequeno aumento nas massas fresca e seca no fruto do café conilon nessa fase (Figuras 6A, B e C), uma vez que o teor de água praticamente não se altera (Figura 6D). Ainda, o teor de açúcares e a respiração da polpa do fruto aumentam durante o amadurecimento (CANNELL, 1971).
5 RELAÇÕES HÍDRICAS
As folhas do cafeeiro mantêm um elevado teor relativo de água, mesmo a potenciais hídricos consideravelmente negativos (DaMATTA et al., 1993, 2002a; PINHEIRO et al., 2005), provavelmente um meio de o café conilon evitar, em vez de tolerar, a dessecação (DaMATTA et al., 1993). Portanto, sintomas visíveis de murcha na folhagem são raros, a menos que a umidade do solo seja muito baixa. O alto teor relativo de água no ponto de perda de turgescência, usualmente próximo ou superior a 90%, parece estar largamente associado à alta rigidez da parede celular. Nesse ponto, os potenciais hídricos situam-se na faixa de -1,7 a -2,2 MPa em clones de café conilon irrigados, com valores mais negativos para clones não irrigados exibindo algum grau de ajustamento osmótico (aumento líquido na concentração de solutos do suco celular), conforme sumariado por DaMatta e Ramalho (2006). No entanto, o ajustamento osmótico, que em algumas espécies é associado com maior produtividade sob condições de seca, não é uma característica geral observada em café. Ademais, a desidratação foliar pode desenvolver-se mais rapidamente após a suspensão da irrigação em clones exibindo maior Figura 6. Variações da massa fresca (A), massa seca (B), taxa de crescimento (C), teor de água (D) e do número de frutos de café por roseta (E), durante a frutificação de clones (48 e 67) de café conilon de maturação precoce. Fonte: Ronchi, DaMatta e Serrano (2006). Nota: A antese ocorreu no dia 2 de setembro de 2005 (Média ± desvio padrão; n = 12; MF = massa fresca). amplitude de ajuste osmótico (DaMATTA, 2004b). Ressalte-se, ainda, que a tolerância à seca, tanto em café conilon como em arábica, não se correlaciona satisfatoriamente com o acúmulo de solutos não tóxicos osmoticamente ativos (e.g., prolina e compostos de amônio quaternário), associado com o ajuste osmótico. Não obstante, o acúmulo desses solutos parece um indicador sensível do status hídrico foliar do cafeeiro (MAESTRI et al., 1995).

Em um experimento com plantas jovens de conilon cultivadas em vasos, DaMatta et al. (2002a, 2002b) observaram que aplicações extras de nitrogênio não afetaram o comportamento estomático, independentemente do status hídrico foliar, mas acarretaram aumentos na rigidez da parede celular e provocaram ajustamento osmótico, quando as plantas foram submetidas à seca. Conjuntamente, esses resultados devem produzir menores potenciais hídricos na planta, para um dado grau de hidratação, o que favoreceria o aumento da eficiência de absorção da água do solo. Resta demonstrar se esses efeitos são também observados com plantas no campo.
6 MOVIMENTO ESTOMÁTICO E TRANSPIRAÇÃO
Comparativamente, o café conilon apresenta maior densidade estomática, mas com estômatos menores que no café arábica os quais são predominantemente do tipo paracítico, enquanto no conilon predomina o tipo actinocítico (CARVALHO et al., 2001). Em ambas as espécies, o fechamento estomático tem sido frequentemente considerado como o indicador primário do deficit hídrico (DaMATTA, 2004a, 2004b). Pinheiro et al. (2005) demonstraram que a condutância estomática (grosso modo, uma medida da abertura do estômato) decresce curso linearmente com a redução da disponibilidade hídrica, sugerindo alta sensibilidade estomática ao desenvolvimento de deficit hídricos na planta de café conilon.
No entanto, esses autores observaram rápida recuperação (1-2 dias) da hidratação foliar e da condutância estomática, que alcançaram valores típicos de cafeeiros irrigados continuamente, após recarregarem clones de conilon severamente desidratados. Isso foi acompanhado da recuperação completa das taxas de fotossíntese (PINHEIRO; DaMATTA, resultados não publicados). Concomitantemente, esse comportamento parece ser de grande importância na determinação do rápido restabelecimento do crescimento e desenvolvimento da copa do café após períodos prolongados de seca e atesta um alto grau de tolerância à seca ao café conilon (DaMATTA; RENA, 2001; DaMATTA; RAMALHO, 2006).
O estômato do café conilon pode responder às variações na demanda evaporativa da atmosfera independentemente do status hídrico foliar (PINHEIRO et al., 2005). De modo geral, clones de conilon mostram alta sensibilidade estomática à demanda evaporativa, mas aparentemente menor que a das cultivares de arábica (DaMATTA, 2004a; DaMATTA; RAMALHO, 2006). Essa menor sensibilidade estomática do conilon pode estar associada a uma maior condutância hidráulica do xilema e maior densidade de venação foliar (SILVA; DaMATTA, resultados não publicados), o que favoreceria uma reposição mais rápida da água perdida pela transpiração, com consequente manutenção, por mais tempo, da abertura estomática, garantindo maiores taxas de influxo de CO2 e, pois, maiores taxas fotossintéticas ao longo do tempo. Isso pode explicar, pelo menos em parte, o porquê de genótipos de conilon responderem melhor à irrigação que os de arábica (DaMATTA, 2004a, 2004b). Salienta-se, contudo, que há clones de conilon (e.g., 109A) que pouco respondem às variações da demanda evaporativa (PINHEIRO et al., 2005) e, por essa razão, sofrem desidratação relativamente rápida em períodos de estiagem, o que concorre para uma baixa tolerância à seca. No entanto, a sensibilidade estomática às variações da umidade relativa do ar aumenta substancialmente no clone 109A, quando enxertado sobre o clone 120, assemelhando-se à dos pés-francos do último. De modo oposto, no clone 120 enxertado sobre o clone 109A, a sensibilidade estomática à umidade relativa diminui (SILVA et al., 2010). Essas respostas estão provavelmente associadas a sinais radiculares (e.g., ácido abscísico) que estariam envolvidos no controle das trocas gasosas da parte aérea (SILVA, LOUREIRO, DaMATTA, resultados não publicados). De qualquer modo, os resultados de SILVA et al. (2010) sugerem que a enxertia, com o uso de porta-enxertos tolerantes à seca, pode ser uma técnica para aumentar a tolerância do café conilon ao deficit hídrico.
A resposta direta do estômato a variações na umidade relativa tem consequências importantes no que respeita à capacidade da planta de suportar períodos relativamente longos de deficit hídrico no solo associados a condições de alta demanda evaporativa na atmosfera. Sob tais condições, o rápido fechamento do estômato seria vantajoso para o cafeeiro, visto que isso resultaria em diminuição substancial da transpiração. Por outro lado, sob disponibilidade hídrica adequada ou sob períodos curtos de deficit hídrico, a sensibilidade do estômato à umidade do ar seria desvantajosa. Nessas condições, a maximização da absorção do CO2 para a fotossíntese passaria a ter maior importância que a redução da transpiração, em termos de manutenção da produtividade, sob condições de seca (DaMATTA, 2003).
Em geral, a condutância estomática é maior em plantas (ou folhas) à sombra que a pleno sol. Os maiores valores de condutância estomática são observados nas primeiras horas da manhã e declinam após as 09:00-10:00 h, podendo alcançar valores negligenciáveis à tarde, associados a altas irradiâncias, temperatura e deficit de pressão de vapor (RODRÍGUEZ-LÓPEZ et al., 2013). Entretanto, em virtude da forte interação entre as respostas dos fatores abióticos mencionados, é muito difícil isolar os seus efeitos. De qualquer modo, aparentemente a temperatura e o deficit de pressão de vapor, mas não a radiação fotossinteticamente ativa, seriam os fatores mais importantes no controle do comportamento estomático do conilon, desde que a irradiância esteja acima de um valor crítico (DaMATTA, 2004b).
7 FOTOSSÍNTESE E PRODUTIVIDADE
As taxas de fotossíntese líquida do café conilon são relativamente baixas (DaMATTA; MAESTRI; BARROS, 1997; DaMATTA et al., 2000; LIMA et al., 2002; DaMATTA, 2003; PINHEIRO et al., 2004, 2005; RONCHI et al., 2005; PRAXEDES et al., 2006; MORAIS et al., 2012b; SILVA et al., 2013) registrando-se, até então, valores máximos em torno de 11-12 µmol CO2 m-2 s-1 para folhas isoladas (PINHEIRO et al., 2004; SILVA et al., 2013). Isso indica que o conilon, assim como as cultivares de café arábica, exibe baixa eficiência fotossintética em relação à da maioria das plantas lenhosas (CANNELL, 1985). Por outro lado, a capacidade fotossintética (fotossíntese potencial), determinada sob luz e CO2 saturantes e à temperatura ótima, alcança valores da ordem de 30 a 35 µmol O2 m-2 s-1 (DaMATTA; MAESTRI; BARROS, 1997; CAMPOSTRINI; MAESTRI, 1998; DaMATTA et al., 2001). Os mecanismos que explicariam os baixos valores das taxas fotossintéticas do cafeeiro estão associados, fundamentalmente, a uma alta resistência à difusão do CO2 , desde a atmosfera até o cloroplasto (tanto em níveis de estômato como de mesofilo) (MARTINS et al., 2014b).
A comparação entre os vários resultados disponíveis na literatura permite inferir que as cultivares de café arábica e clones de conilon exibem taxas máximas de fotossíntese líquida similares. No entanto, em face da provável maior sensibilidade estomática à demanda evaporativa do ar em arábica que em conilon, é lícito sugerir que o último possa manter, ao longo do dia, maior absorção cumulativa de CO2 . Ademais, o conilon parece capaz de manter taxas fotossintéticas relativamente muito mais elevadas na época de baixo crescimento vegetativo (abril a setembro), quando comparadas às do café arábica (SILVA et al., 2004; MORAIS et al., 2012b). Por conseguinte, maiores taxas globais de fotossíntese, tanto em escala diária como em escala anual, poderiam refletir-se na maior produção de frutos em conilon que em arábica, conforme frequentemente observado. Ademais, o café conilon é capaz de acumular grandes concentrações foliares de amido (~14% em base massa seca) sem qualquer retroinibição metabólica aparente da fotossíntese (MORAIS et al., 2012b).
Considerando-se que 90-95% da massa seca das plantas se derivam da fotossíntese, não é difícil perceber que a produção depende, direta ou indiretamente, da magnitude das taxas fotossintéticas (KHANNA-CHOPRA, 2000). Com efeito, clones de maturação precoce, que têm uma maior demanda por assimilados dentro de um prazo mais curto (maior demanda dos drenos), exibem maiores taxas fotossintéticas acopladas a maiores condutâncias estomáticas (MORAIS et al., 2012b). Coerentemente, Partelli et al. (2014c) constataram que clones de maturação precoce exibem maior taxa de acúmulo de matéria seca de frutos, assim como de nutrientes, em comparação a clones de maturação mais tardia. Os mecanismos fisiológicos pelos quais aumentos na capacidade fotossintética estão associados com o padrão de maturação de frutos são completamente desconhecidos. Em todo o caso, a existência de um padrão de regulação da fonte (capacidade fotossintética das folhas) pela maior demanda dos drenos (frutos) pode abrir novas oportunidades dentro das fronteiras da fisiologia molecular para verificarem-se genes e fatores de transcrição que poderiam auxiliar no aumento da compreensão dos mecanismos que governam as relações fonte/ dreno em café. Não obstante, é pouco provável que diferenças clonais de produtividade estejam fundamentalmente associadas com potenciais variações nas taxas de fotossíntese líquida de folhas isoladas (CAMPOSTRINI; MAESTRI, 1998; DaMATTA et al., 2000). Provavelmente, essas diferenças estariam associadas mais diretamente com a manutenção de uma área foliar sadia (maiores taxas de fotossíntese da planta inteira, ao longo do tempo) e com uma arquitetura de copa mais favorável à maximização das trocas gasosas. Além disso, alterações nas condições de crescimento podem promover alterações não somente na magnitude das taxas de fotossíntese, mas também na partição de assimilados entre órgãos vegetativos e reprodutivos, modificando, portanto, o índice de colheita, aspectos esses virtualmente inexplorados em café.
8 ESTRESSE OXIDATIVO
No metabolismo celular, uma consequência inevitável das reações de transporte de elétrons do cloroplasto é a transferência de elétrons para o oxigênio molecular levando à formação de espécies reativas de oxigênio. No aparelho fotossintético, isso pode causar estresse oxidativo, fenômeno comum promovido por vários tipos de estresses (e.g., seca, salinidade, extremos de temperatura, adubação deficiente), que geram um decréscimo no consumo de energia sem uma redução significante na sua absorção (DaMATTA; RAMALHO, 2006). Proteção contra o estresse oxidativo pode ocorrer a expensas de aumento na dissipação da energia de excitação por meio de carotenoides ou via metabolismo das espécies reativas de oxigênio, mediante maior atividade do sistema antioxidativo. Este, por sua vez, envolve moléculas de massas variáveis (e.g., β-caroteno, ascorbato, glutationa) e enzimas antioxidativas (e.g., dismutase do superóxido, peroxidases etc.).
No cafeeiro, como na maioria das plantas C3 , a fotossíntese é saturada a cerca de 1/3 da irradiância máxima incidente sobre a copa, i.e., cerca de 550-700 µmol (fótons) m-2 s-1 em folhas expostas (DaMATTA; MAESTRI, 1997; RODRÍGUEZ-LÓPEZ et al., 2013). A irradiância excedente absorvida pela folha, se não dissipada satisfatoriamente, pode acarretar perda do controle do metabolismo das espécies reativas de oxigênio, levando à ocorrência de danos fotooxidativos (DaMATTA; RAMALHO, 2006), popularmente conhecidos como escaldadura. Esses danos se manifestam pelo aparecimento de áreas cloróticas, que evoluem para a necrose, levando, pois, à abscisão foliar.
A pleno sol, cafeeiros bem adubados e irrigados devem dissipar satisfatoriamente o excesso de energia radiante que recebem (RODRÍGUEZLÓPEZ et al., 2013) e, portanto, a escaldadura, caso ocorrente, deve afetar apenas marginalmente a produção do cafezal. Isso explicaria, pelo menos em parte, o sucesso do cultivo do conilon, como também das cultivares arábicas a pleno sol (DaMATTA; MAESTRI, 1997), ainda que as espécies sejam originalmente de ambientes sombreados. Por outro lado, sob certas condições, os mecanismos de defesa da planta podem não ser suficientes para dissipar a energia excedente e, nesse caso, a ocorrência de escaldadura pode ser bastante expressiva. Entre tais condições, destacam-se: cafezais mal-adubados, carga grande de frutos, deficit hídrico e extremos de temperatura ou qualquer outro motivo que leve ao depauperamento da planta. Esses fatores, combinados ou não, geralmente resultam no aparecimento da escaldadura, culminando com a abscisão da folha. Nessas condições, a incidência de bicho-mineiro, cercóspora e ferrugem usualmente aumenta, também levando à abscisão foliar. Desse modo, não somente a folhagem exposta mas também as folhas mais internas da copa passam a receber irradiância superior à necessária para saturar a fotossíntese, o que potencializa mais ainda a abscisão das folhas. Em muitos casos, o ramo seca ou pouco cresce, acarretando quebra de produção na colheita seguinte. O sombreamento, nesse contexto, poderia minimizar, sobremodo, a ocorrência da escaldadura, especialmente se associado à irrigação (DaMATTA; RENA, 2002).
Lima et al. (2002), avaliando dois clones de café conilon sob condições de seca, observaram aumentos muito maiores na atividade do sistema antioxidante do clone 120 (tolerante à seca) que no clone 109A (sensível à seca). Os autores também mostraram que a extensão de danos celulares foi substancialmente menor no clone tolerante que no sensível à seca, o que os levou a propor que a tolerância à seca no conilon poderia, pelo menos em parte, estar associada a um incremento da atividade do sistema antioxidante. Resultados de Pinheiro et al. (2004) indicaram que o clone 120, capaz de tolerar períodos de seca relativamente longos com produção aceitável nessa condição, demonstrou maior capacidade de tolerar o estresse oxidativo induzido pelo herbicida paraquat que outros clones avaliados (14, 46, 109A). Esses dados foram reforçados pelos resultados de Menezes (2005), que observou que a atividade de uma enzima do sistema antioxidante (dismutase do superóxido) foi máxima a concentrações de paraquat da ordem de 20 µM, enquanto no clone 109A a atividade da enzima foi virtualmente nula. Diante dessas informações, DaMatta e Ramalho (2006) propuseram que um sistema antioxidante mais eficiente pode ter uma função decisiva na tolerância à seca em café, particularmente em clones de conilon que favorecem a produção em detrimento da sobrevivência. Sistemas antioxidantes mais robustos permitiriam, pois, maior tolerância protoplasmática à dessecação.
Em condições de altas temperaturas e baixa umidade relativa, o sombreamento do café conilon pode minimizar os efeitos deletérios da alta demanda evaporativa da atmosfera (PEZZOPANE et al., 2011; PARTELLI et al., 2014a) e, desse modo, promover melhorias no desempenho fotossintético e reduzir potenciais danos oxidativos (RODRÍGUEZ-LÓPEZ et al., 2013). Não obstante, parece haver variabilidade clonal no que respeita às respostas fotossintéticas à disponibilidade de luz (RODRÍGUEZ-LÓPEZ et al., 2013). Pesquisas são necessárias, portanto, para explorar essa variabilidade, com vistas à indicação de clones mais promissores para serem utilizados em sistemas agroflorestais (sombreados).
9 TOLERÂNCIA À SECA
No Brasil, e especialmente no Espírito Santo, o cultivo do café conilon tem-se expandido para áreas onde a deficiência hídrica é o principal fator limitante à produção. Para melhorar o rendimento de cafeeiros em ambientes com restrição hídrica, melhoristas têm procurado selecionar genótipos que produzam razoavelmente bem nesses ambientes. Isso nem sempre é tarefa fácil, em razão da variabilidade da precipitação ao longo dos anos e da natureza poligênica da tolerância à seca (DaMATTA, 2004b). Uma abordagem complementar para melhorar o desempenho do conilon em regiões sujeitas à seca residiria na seleção de índices e parâmetros adequados que permitam identificar materiais genéticos com maior potencial de tolerância à seca. Estudos comparativos, tanto com plantas em vasos como no campo, têm evidenciado que, quando submetidos à deficiência hídrica, os clones tolerantes são capazes de manter melhor hidratação (potencial hídrico menos negativo) de seus tecidos que os clones sensíveis à seca (DaMATTA et al., 2000, 2003; PINHEIRO et al., 2004, 2005; PRAXEDES et al., 2006; SILVA et al., 2013). Essa estratégia explicaria o porquê de os clones tolerantes exibirem enrolamento e abscisão foliar mais tardiamente que os clones sensíveis à seca.
Ao longo de períodos prolongados de estiagem, contudo, melhor hidratação tem sido associada, em muitos casos, com estratégias de sobrevivência, e não com manutenção da produção sob condições de seca. Nesse contexto, o fechamento estomático eficiente, conforme observado em clones mais tolerantes à seca, parece estar associado a uma rede complexa de respostas envolvendo rotas de sinalização mediadas pelo ácido abscísico e pelo óxido nítrico (MARRACCINI et al., 2012). Em todo o caso, um fechamento estomático eficiente para limitar a transpiração concorre, ao mesmo tempo, para limitar o influxo de CO2 para a fotossíntese e, portanto, a produção. Por essa mesma razão, pouco adianta, também, manter uma área foliar sadia e, assim, o vigor, se esse comportamento estiver associado à restrição estomática às trocas gasosas da folhagem. Registra-se, não obstante, que a manutenção da área foliar pouparia recursos à planta que, de outro modo, teriam de ser despendidos para a recuperação da área foliar após períodos de estiagem (DaMATTA; RAMALHO, 2006).
Estudos conduzidos em Sooretama/ES têm mostrado grande diversidade genética entre clones de conilon no que respeita à produção sob condições de seca (FERRÃO et al., 2000a; 2000b). Nessa condição, observam-se clones sensíveis à seca, com baixo vigor, desfolha considerável e grande comprometimento da produção; e clones que exibem bom vigor, mas sem manter necessariamente produção relativamente elevada sob seca. Diferenças morfofisiológicas entre esses clones devem, portanto, responder grandemente por suas tolerâncias diferenciais à seca (DaMATTA; RAMALHO, 2006). A partir dos resultados de DaMatta et al. (2000, 2003), Pinheiro et al. (2004, 2005) e de Praxedes et al. (2006), sugerem-se três estratégias possivelmente envolvidas nessa tolerância diferencial:
(i) os clones sensíveis à seca mantêm um controle deficiente da transpiração (clone 109A), com seus estômatos respondendo de forma limitada à redução da disponibilidade de água do solo ou à demanda evaporativa da atmosfera. Alternativamente, ainda que exibam controle estomático satisfatório da transpiração, podem ter baixa capacidade de absorção da água, visto que têm sistemas radiculares mais superficiais (clone 46). Em quaisquer desses casos, a desidratação dos tecidos é acelerada sob deficiência hídrica, levando ao colapso do metabolismo, culminando com perda considerável da área foliar e, em última instância, comprometendo irremediavelmente a produção. A exploração econômica desses clones requer, via de regra, a implementação de irrigação;
(ii) clones tolerantes com sistema radicular relativamente profundo, alta sensibilidade estomática à disponibilidade de água e baixas taxas do uso da água (baixa condutância hidráulica) postergam a desidratação de seus tecidos, mantendo um balanço hídrico favorável e, assim, o vigor. Não obstante, ao mesmo tempo que os estômatos se fecham para limitar a transpiração, restringem também o influxo de CO2 para a fotossíntese e, por extensão, a produção é reduzida (clone 14). Clones com esse comportamento normalmente respondem de forma insatisfatória à irrigação se a demanda evaporativa da região for alta, isto é, ainda que haja disponibilidade adequada de água no solo, seus estômatos se fecham em resposta à baixa umidade relativa do ar, comprometendo a produção. Essa estratégia de tolerância à seca favorece a sobrevivência em detrimento da produção;
(iii) clones tolerantes com sistemas radiculares relativamente profundos e sensibilidade estomática satisfatória à disponibilidade de água, mas com alta condutância hidráulica, têm comportamento intermediário no que concerne à taxa de desidratação de seus tecidos sob seca (clone 120). Com disponibilidade adequada de água no solo, esses clones aliam taxas de transpiração relativamente elevadas (e, portanto, maior influxo de CO2 para a fotossíntese) com rápida reposição da água transpirada. À medida que a disponibilidade de água do solo diminui, o sistema radicular relativamente profundo garantiria a manutenção parcial da absorção da água; no entanto, isso faz exaurir mais rapidamente as reservas de água do solo, concorrendo para o desenvolvimento de deficit hídricos internos. Esses clones, possivelmente, têm certo grau de tolerância protoplasmática (sistema antioxidante mais eficiente) à dessecação, o que preveniria o colapso do metabolismo celular. Normalmente, clones com esse comportamento produzem satisfatoriamente tanto em condições de seca como irrigadas, sendo, pois, denominados de clones de dupla aptidão.
Em suma, os principais componentes da adaptação diferencial à seca entre clones de conilon parecem ser comportamentais, sendo provavelmente governados pelas taxas de uso da água e/ou pela eficiência de extração da água do solo (DaMATTA et al., 2000, 2003; PINHEIRO et al., 2005; DaMATTA; RAMALHO, 2006), ou pela capacidade de condução do xilema (SILVA et al., 2013). De um ponto de vista molecular, diferentes mecanismos parecem envolvidos na tolerância diferencial à seca em conilon envolvendo rotas de sinalização dependentes e independentes do ácido abscísico (MARRACCINI et al., 2012; VIEIRA et al., 2013). Em todo o caso, avaliações fisiológicas sugerem que clones com produção relativamente elevada sob seca o fazem via manutenção de status hídrico adequado (via combinação de sistemas radiculares profundos e controle estomático satisfatório da transpiração) e manutenção da área foliar com folhas com orientação mais verticalizada. Além disso, características bioquímicas seriam também importantes, como a existência de sistema antioxidativo mais robusto e manutenção da capacidade de exportação (o que permitiria, por exemplo, o crescimento radicular e, assim, maior acesso a horizontes mais profundos do solo, de modo a aumentar a absorção de água). Essas características parecem não assumir o mesmo papel de destaque nos clones que favorecem a sobrevivência em detrimento da produção. Como um todo, essas informações sugerem a dificuldade de se estabelecer uma característica isolada capaz de predizer adequadamente a tolerância à seca do conilon, em termos de manutenção da produção em ambientes sujeitos à restrição hídrica (DaMATTA; RAMALHO, 2006).
A despeito das considerações acima, Silva et al. (2013), ao explorarem a divergência funcional entre diversos clones de conilon, em termos de ganho de carbono e uso da água, demonstraram que clones com baixa densidade do caule apresentam maiores taxas de uso de água e ganho de carbono sob disponibilidade hídrica adequada. Sob seca, contudo, esses clones utilizam mais rapidamente a água disponível, o que gera sintomas mais precoces da seca, menor taxa fotossintética e menor acúmulo de biomassa. Esses autores sugeriram que a densidade do caule, um parâmetro de fácil mensuração, poderia ser usada para predizer a tolerância à seca em conilon, além de ser um integrador adequado das dicotomias existentes entre segurança e eficiência hidráulica do xilema no cafeeiro. Em outras palavras, clones com menor densidade do caule possuiriam uma maior capacidade de condução de seiva e, assim, seriam capazes de suportar maiores taxas de trocas gasosas e acúmulo de biomassa sob hidratação adequada. Sob seca, esses clones sofrem mais facilmente cavitação e embolia, situação que resulta em colapso hidráulico e menor capacidade de acúmulo de biomassa e sobrevivência. Enfatizase, não obstante, que tolerância à seca associada à conservação de água pode ser uma estratégia mais eficiente para sobrevivência do que para manutenção, dentro de certos limites, da produção de frutos.
Outro aspecto importante que vem sendo atualmente avaliado reside na análise fisiológica da tolerância à seca do cafeeiro na fase de muda, e se uma presumível tolerância na muda seria conservada na planta adulta, no campo. Os primeiros resultados parecem promissores. Se confirmados, seria possível a discriminação precoce de genótipos com maior potencial de tolerância à seca resultando em economia de tempo e otimização de recursos, porquanto se permitiria o descarte precoce de genótipos menos promissores, ou, de modo oposto, permitindo a concentração de esforços em materiais potencialmente mais produtivos, sobre os quais, em última instância, baseia-se um programa de melhoramento (CAVATTE et al., 2008; SILVA et al., 2013).
10 DIFICULDADES E AVANÇOS PARA A OBTENÇÃO DE CULTIVARES TOLERANTES À SECA
De modo geral, os progressos observados nos últimos 100 anos, no que respeita à obtenção de cultivares superiores em termos de tolerância à seca, têm sido muito tênues na grande maioria dos programas de melhoramento, independentemente da cultura explorada, diferentemente do que ocorre com o melhoramento visando à tolerância contra estresses bióticos e outros estresses abióticos (CAVATTE et al., 2012).
Faz-se necessário, primeiramente, definir o que é um genótipo tolerante à seca. Usualmente, os melhoristas centram a sua atenção em genótipos que produzam mais que outros, quando submetidos ao deficit hídrico. Todavia, essa definição pode não ser a mais adequada. Considerando-se, por exemplo, dois clones de café conilon, A e B, que produzissem, respectivamente, 100 e 60 sc ha-1 sob irrigação, e 50 e 40 sc ha-1 num ano seco, sem irrigação, perguntase: qual deles seria o mais tolerante à seca? Certamente, o clone B, cuja produção teria sido reduzida em 33%, contra 50% no clone A, apesar da maior produção absoluta deste clone (porém, normalmente às expensas de maiores custos de produção). Em outras palavras, o clone B seria menos afetado pelo deficit hídrico e, portanto, seria mais tolerante à seca. Genótipos mais tolerantes à seca têm, normalmente, maior estabilidade de produção (mas geralmente com menor potencial produtivo), podendo ser recomendados para sistemas de produção com menor nível tecnológico, apesar de os clones de café conilon de aptidão dupla produzirem satisfatoriamente tanto com ou sem irrigação.
Enfatiza-se que a tolerância à seca é uma resultante de várias características (anatômicas, morfológicas, fisiológicas, moleculares), que se expressam diferente e concomitantemente, dependendo da severidade e da taxa de imposição do deficit hídrico, da idade e das condições nutricionais da planta, do tipo e da profundidade do solo, da carga pendente de frutos, da demanda evaporativa da atmosfera, da face de exposição do terreno etc (DaMATTA, 2003). Portanto, a adoção de uma única estratégia de adaptação à seca é, certamente, inadequada para qualquer tipo de ambiente. De um ponto de vista fisiológico, muitos experimentos têm sido conduzidos com plantas em pequenos vasos, em casas de vegetação ou em salas de crescimento, submetendo-se as plantas a secas drásticas e rapidamente impostas. As respostas assim obtidas guardam pouco paralelismo com o que se observa no campo e, portanto, os resultados comparativos auferidos devem ser analisados com reserva. Além do mais, em muitos casos, as análises têm sido feitas em genótipos com diferentes graus de hidratação e, pois, não são rigorosamente comparáveis. Consequentemente, boa parte das informações geradas por fisiologistas, no que concerne à tolerância à seca, tem utilidade limitada para o melhorista, porquanto tais informações não podem ser convertidas em índices adequados para a predição do comportamento de genótipos com tolerância diferencial ao deficit hídrico. A dissociação entre as informações básicas e as necessidades de informações aplicadas (ou aplicáveis) constituiemse num gargalo adicional para avançar-se na obtenção de materiais mais tolerantes à seca (CAVATTE et al., 2012).
Para o café conilon, apesar de não se ter nenhuma análise concreta da fração do ganho de produtividade associada com a tolerância per se à seca (ou ao manejo em si) de materiais genéticos de destaque, é lícito presumir-se que o Programa de Melhoramento do Incaper vem explorando adequadamente a variabilidade genética existente nas populações de café conilon. Isso parece bem ilustrado (i) com a cultivar clonal ‘Vitória Incaper 8142’, lançada em 2004, com produção média, em ensaios não irrigados, de 70,40 sc ha-1, contra uma produção média capixaba de 24,12 sc ha-1 à época (FONSECA et. al., 2004; FERRÃO et al., 2007); e (ii) com o lançamento, em 2013, de três novas cultivares clonais (‘Diamante ES8112’, ‘ES8122 - Jequitibá’ e ‘Centenária ES8132’) também com tolerância à seca, com produtividades superiores a 80 sc ha-1 (FERRÃO et al., 2015a, 2015b, 2015c). Em todo o caso, devido ao estreitamento da base genética da espécie, com a adoção de cultivares clonais, os melhoristas têm que, continuamente, procurar desenvolver novos materiais genéticos com alto potencial produtivo, mesmo sob condições de seca. Espera-se que a utilização de critérios fisiológicos para auxiliar na identificação de genótipos superiores sob deficit hídrico, aliados a critérios fitotécnicos, facilitará sobremodo essa tarefa.
11 MUDANÇAS CLIMÁTICAS E O FUTURO DO CONILON
O aumento da temperatura e da concentração atmosférica de CO2 (), associado às variações nos padrões de precipitação, constitui, em grande parte, o escopo das chamadas mudanças climáticas globais (DaMATTA et al., 2010). Dada a sensibilidade do cafeeiro às altas temperaturas, especialmente o café arábica, extensas áreas ocupadas pela cafeicultura, hoje consideradas aptas, passarão a ser marginais ou inaptas, como tem sido sugerido em estudos meteorológicos de simulação (ZULLO et al.,2011). Em razão das exigências diferenciais relativas em termos de temperatura, entre o café arábica e o conilon, é mais que óbvio especular-se que o comportamento dessas espécies deverá variar em resposta às mudanças climáticas. Assim, algumas áreas hoje marginais ao café conilon, notadamente áreas do Sudeste do Brasil em altitudes superiores a 500 m, poderão ter parte de suas lavouras de arábica substituídas por conilon.
Uma vez que a taxa fotossintética do cafeeiro é grandemente limitada por restrições difusivas, e não por restrições bioquímicas (MARTINS et al., 2014a), sugere-se que o conilon tenha um grande potencial para aumentar o seu desempenho fotossintético com o aumento da . Portanto, a quantidade de CO2 sequestrada pelo cafeeiro deve aumentar, em termos relativos, muito mais do que se prevê para várias outras espécies perenes (MARTINS et al., 2014a), como o eucalipto. Com efeito, em experimentos de longa duração com café conilon (clone 153) cultivado em vasos, em câmaras de crescimento, têm-se observado maiores taxas de fotossíntese (49% maiores a uma = 700 ppm em relação à = 380 ppm) associadas com invariabilidade da condutância estomática, resultando em um excepcional incremento na eficiência do uso da água (RAMALHO et al., 2013). Por conseguinte, o aumento da provavelmente possibilitará ao cafeeiro produzir mais assimilados (e mais frutos) e usar menos água na ausência de outros estresses. Registra-se, ainda, que o aumento da temperatura per se impacta negativamente o desempenho fotossintético do conilon, mas esses efeitos podem ser largamente mitigados pelo aumento da (RAMALHO et al., 2014). Outro aspecto a se considerar reside na menor concentração de nutrientes (N, Mg, Ca e Fe) do conilon cultivado em alta (700 ppm) em relação à ambiente (380 ppm), mas tais decréscimos podem ser atenuados ou anulados pelo aumento da temperatura (MARTINS et al., 2014b), provavelmente um reflexo da menor expansão foliar, de modo a reverter os efeitos de diluição de nutrientes observados em decorrência da alta .
Grandes incertezas permanecem, contudo, sobre a produção e a qualidade da bebida em um cenário envolvendo incremento da , aquecimento global e alteração nos padrões de precipitação. Por um lado, os aumentos esperados nas taxas fotossintéticas associadas com o incremento da implicarão numa melhor capacidade de enchimento do grão, o que pode resultar em melhor qualidade da bebida. Por outro lado, os impactos benéficos diretos do CO2 poderão ser reduzidos ou mesmo anulados por alterações em outros fatores do ambiente, como a elevada temperatura, que pode comprometer a produção e acelerar o crescimento e a maturação dos frutos, reduzindo a qualidade da bebida (DaMATTA; CAVATTE; MARTINS, 2012). Essas limitações e outras mudanças climáticas, como secas mais frequentes e severas, podem adicionalmente produzir novas condições limitantes para o crescimento do cafeeiro. Em conjunto, esses fatos bem ilustram a alta complexidade envolvida na previsão do impacto do aumento da e temperatura e outras mudanças climáticas ora em curso sobre a produção e qualidade do café.
12 CONSIDERAÇÕES FINAIS
O conhecimento sobre a fisiologia do café conilon é incipiente, se comparado ao do café arábica. Nesse contexto, pouco se tem explorado sobre o crescimento vegetativo e, muito menos ainda, sobre o crescimento reprodutivo (floração e frutificação) do café conilon nos vários ambientes e sistemas de cultivo. As poucas informações disponíveis resumem- -se a aspectos observacionais, sem a exploração pormenorizada dos mecanismos fisiológicos que explicariam aqueles processos. Pouco se conhece sobre a fisiologia do sistema radicular, apesar de alguns avanços terem ocorrido na elucidação de sua morfologia e distribuição no solo. Ademais, estudos sobre os componentes fisiológicos da produção são incipientes. Alguns aspectos sobre a fotossíntese e as relações hídricas têm sido explorados com algum grau de aprofundamento, mas as relações hormonais, o metabolismo do nitrogênio e de outros minerais, a partição de assimilados, entre outros, não têm sido estudados. O uso de ferramentas de biologia molecular para avançar-se na compreensão da fisiologia do café conilon é ainda embrionário e restrito a muito poucos laboratórios. Em resumo, as inúmeras deficiências no conhecimento sobre a fisiologia do café conilon têm produzido um campo fértil de desafios a serem vencidos em pesquisas vindouras.
Em um país com tantas carências de recursos humanos e financeiros para pesquisas básicas, e mesmo para aplicadas, espera-se que os esforços em investigação científica se voltem principalmente para aumentar a base de conhecimento sobre as interações fisiológicas do café conilon com o seu ambiente, de modo a subsidiar a racionalização do manejo da lavoura cafeeira em diferentes sistemas de produção.
Fonte
Cláudio Pagotto Ronchi e Fábio Murilo DaMatta; Aspectos Fisiológicos do Café Conilon; In: FERRÃO, R. G., DE MUNER, L. H., da FONSECA, A. F. A., & Ferrão, M. A. G. (2016). Café Conilon. Vitória, ES: Incaper, 2017..



